Образец резюме специалиста без опыта работы на желаемой должности
Добавить фото
Иванов Иван
Пол, дата рождения
Контактный телефон
Контактный эл. адрес
Проживает: город
Гражданство:
Готов \ не готов к переезду, готов \ не готов к командировкам
Желаемая должность и зарплата
Специалист по организации мероприятий
Маркетинг, реклама, PR
- Продвижение, специальные мероприятия
Занятость: полная занятость
График работы: полный день
Желательное время в пути до работы: не имеет значения
Когда опыта работы по специальности нет, постарайтесь раскрыть свои преимущества в сопроводительном письме. По структуре оно может быть таким:- Почему вы хотите работать именно в этой компании.

- Почему вы хотите работать на этой должности. Что уже умеете и как это поможет в новой профессии, что хотите развить и какой опыт получить.

Опыт работы — 3 года 11 месяцев
Октябрь 2016 — настоящее время1 год 1 месяц
Октябрь 2016 — настоящее время1 год 1 месяц
Москва
Продавец-консультант
Основные обязанности:
- Организация работы с клиентами — физическими лицами.
- Консультирование по продуктам и услугам.
- Выполнение поставленных планов продаж.
- Соблюдение стандартов обслуживания и мерчандайзинга.
- Участие в организации корпоративных мероприятий.

Достижения:
- Выполнял поставленные цели по планам продаж, неоднократно был признан лучшим продавцом месяца.
- Участвовал в организации новогоднего корпоративного мероприятия: разработка концепции, поиск площадки для проведения мероприятия, ведение мероприятия в качестве ведущего. Мероприятие получило высокую оценку коллег и руководителей.
Даже если вы проработали недолго, добавьте все обязанности и задачи, с которыми успели столкнуться хотя бы раз. Это покажет, что хотя бы минимальный опыт у вас есть.
Когда опыта работы по специальности нет, постарайтесь раскрыть свои преимущества в сопроводительном письме. По структуре оно может быть таким:- Почему вы хотите работать именно в этой компании. Ищите что-то общее: например, компания ведет активную социальную поддержку в регионе, а вы тоже участвуете в социальных проектах.
- Почему выхотите работать на этой должности. Что уже умеете и как это поможет в новой профессии, что хотите развить и какой опыт получить.
 Ищите что-то общее: например, компания ведет активную социальную поддержку в регионе, а вы тоже участвуете в социальных проектах.
Ищите что-то общее: например, компания ведет активную социальную поддержку в регионе, а вы тоже участвуете в социальных проектах.Даже если вы проработали недолго, добавьте все обязанности и задачи, с которыми успели столкнуться хотя бы раз. Это покажет, что хотя бы минимальный опыт у вас есть.
Объединяем весь опыт стажировок. Начало периода — начало первой стажировки. Конец периода — окончание последней. В блоке «Обязанности» перечисляем кратко периоды стажировок и выполняемые функции.
Указать, какие конкретно мероприятия.
Можно более подробно описать, почему интересна эта сфера.
Июнь 2014 — Сентябрь 20162 года 10 месяцев
Июнь 2014 — Сентябрь 20162 года 10 месяцев
Стажировки в различных компаниях,
Москва
Стажер
06.
Участие в проекте по подготовке международной фармацевтической конференции:
- взаимодействие с участниками, решение организационных вопросов;
- регистрация участников на мероприятии, консультационно-информационная поддержка.
06.2015 — 07.2015 Производственная компания
Участие в проведении маркетингового исследования возможных потребителей продукции компании:
- поиск информации в сети Интернет и специальных базах;
- заполнение специальных форм отчетности;
- обработка данных с использованием программного обеспечения.
07.2014 — 09.2014 Производственная компания
Административная поддержка работы отдела рекламы:
- работа с документами;
- формирование архива договоров.
Объединяем весь опыт стажировок. Начало периода — начало первой стажировки. Конец периода — окончание последней. В блоке «Обязанности» перечисляем кратко периоды стажировок и выполняемые функции.
Начало периода — начало первой стажировки. Конец периода — окончание последней. В блоке «Обязанности» перечисляем кратко периоды стажировок и выполняемые функции.
Ключевые навыки
Организация мероприятий Управление командой Делопроизводство Организаторские навыки Английский язык Работа в команде Организация встреч Организация деловых поездок Работа на выставках Деловая этика Деловая коммуникация
Рекомендации
по запросу
Обо мне
Опыт организации различных мероприятий: деловых и личных.
В период учебы в вузе участвовал в работе студенческого совета, организовывал различные мероприятия для студентов: спортивные, социальные, творческие.
Опыт участия в проекте по организации международной конференции.

Развитые организационные, коммуникативные и презентационные навыки.
Навыки продаж и ведения переговоров.
Знание основ документооборота и договорной работы.
Опыт проведения маркетинговых исследований рынка.
Умение работать в режиме многозадачности.
Умение работать в команде.
Ориентация на результат.
Желание и готовность развиваться в сфере event-менеджмента.
Уверенный пользователь ПК: MS Office (Word, Excel, Power Point).
Указать, какие конкретно мероприятия.
Можно более подробно описать, почему интересна эта сфера.
Высшее образование
Основное образование: высшее
Дата окончания вуза
Полное наименование вуза
Факультет \ специальность
Знание языков
Русский — родной
Английский — уровень владения языком
Повышение квалификации, курсы
Event-менеджмент
Учебный центр
Как составить резюме, которое заинтересует работодателя
Опыт — первое, на что смотрят работодатели при поиске сотрудника.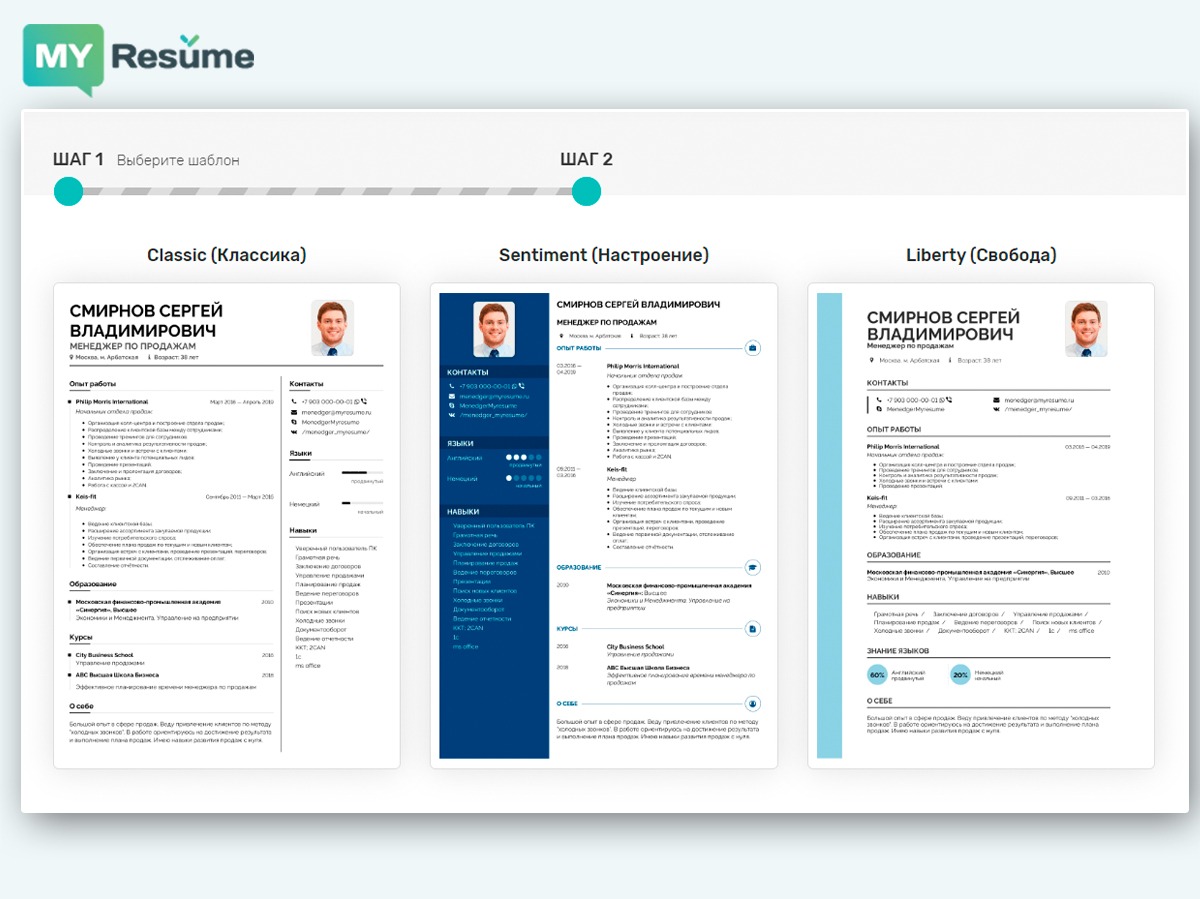 Чтобы произвести положительное впечатление, следуйте простым рекомендациям.
Чтобы произвести положительное впечатление, следуйте простым рекомендациям.
Частая смена работы или длительные пробелы в стаже — тревожный сигнал для руководителя. Если перерывы были, объясните их причины: взяли декретный отпуск, перешли на фриланс, открыли свое дело. Представьте пробел в стаже ценным этапом, который помог вам двигаться дальше.
Начало карьеры мало рассказывает о вашем профессиональном уровне сейчас. Требования к профессиям меняются, нормативы и документы — тоже. Укажите в резюме последние места работы и свои обязанности. Выделите 4-5 основных задач и опишите их понятным менеджеру языком. Начальные этапы карьеры можно упомянуть совсем кратко или соединить несколько записей в одну, если вы часто меняли работу, оставаясь в одной должности.
Говорите о своем опыте в конкретных и измеримых показателях. Что изменилось в компании благодаря вашей работе?
«Занимался оптимизацией бизнес-процессов» — «Разработал корпоративные стандарты контроля качества обслуживания»
«Осуществлял холодные звонки и активные продажи» — «За полгода увеличил базу первичных клиентов компании на 20%»
Пометка о желаемой зарплате отсеет неподходящие вакансии и сориентирует работодателей. Боитесь отпугнуть рекрутера или упустить возможную выгоду? Посмотрите похожие вакансии, сверьте свои пожелания со средней зарплатой по рынку в вашем регионе. Подробнее об этом читайте в одной из наших статей.
Боитесь отпугнуть рекрутера или упустить возможную выгоду? Посмотрите похожие вакансии, сверьте свои пожелания со средней зарплатой по рынку в вашем регионе. Подробнее об этом читайте в одной из наших статей.
Ключевые навыки — специальные знания и умения, которые применяются в конкретной сфере. Для маркетолога подойдут «SWOT-анализ» и «Разработка стратегии продвижения», для финансиста — «Бюджетирование» и «Оценка рисков», и так далее.
Система hh.ru подскажет наиболее короткий и употребимый вариант того или иного навыка — по возможности выбирайте именно его. Избегайте очевидного: знание Windows и Chrome можно не упоминать, а вот навыки «1С: Битрикс» или «b2b-продажи» — ваше преимущество перед другими кандидатами.
Грамотно оформленный раздел «Обо мне» выделяет кандидата и повышает шансы получить приглашение. Пишите только о том, что будет интересно потенциальному работодателю и поможет получить желаемую должность.
Отойдите от клише: коммуникабельность, исполнительность, стрессоустойчивость встречаются в каждом втором резюме. Вспомните свои отличительные черты и методы работы либо опишите привычные качества более развернуто.
Вспомните свои отличительные черты и методы работы либо опишите привычные качества более развернуто.
Плохо: Меня отличает ответственность и пунктуальность.
Лучше: Добросовестно отношусь к выполнению задач. Умею работать в условиях неопределенности и самостоятельно принимать решения.
Выиграли чемпионат города по шахматам или олимпиаду по программированию? Напишите об этом, а общие интересы опустите.
Плохо: Увлекаюсь чтением и спортом.
Лучше: КМС по плаванию и скалолазанию. В свободное время читаю литературу по бизнесу и саморазвитию.
Чтобы обрабатывать резюме быстрее, менеджеры ищут по ключевым словам. Для разработчика это названия языков программирования, для бухгалтера — версии конфигураций. Используйте правильные ключи, чтобы вас было легче найти.
Указывайте простой электронный адрес: в идеале — первая буква имени и фамилия. Это правило делового этикета и признак профессионального подхода к работе. Если нужно, оставьте телефон для связи — одного номера будет достаточно. В строке домашнего адреса укажите город и при желании — ближайшее метро.
Если нужно, оставьте телефон для связи — одного номера будет достаточно. В строке домашнего адреса укажите город и при желании — ближайшее метро.
Фотография в резюме — это плюс, независимо от сферы деятельности и должности. Подойдет портрет, где вы спокойно смотрите в камеру и хорошо видно лицо. В резюме используйте деловое или нейтральное фото, а снимки с отпуска и праздников сохраните для домашнего архива.
Итоговый текст резюме должен представлять собой выжимку, в которой содержится только самая необходимая информация. Большинство резюме однотипны, поэтому лучше писать о себе просто и по делу. Избегайте сложных оборотов и казенных формулировок.
Плохо: Эффективно исполняемые мною обязанности высоко ценились на предыдущем месте работы.
Лучше: Я соблюдаю сроки и аккуратно веду отчетность.
Грамотно написанный текст производит хорошее впечатление, а резюме с ошибками выдает спешку или невнимательность. После заполнения анкеты пройдитесь по тексту, исправьте недочеты и уберите лишнее. Помните: у каждой профессии есть свои особенности. Если от юриста ждут резюме без единой ошибки, то бухгалтерам важнее указать конкретные счета и версии 1С, с которыми они работали.
Помните: у каждой профессии есть свои особенности. Если от юриста ждут резюме без единой ошибки, то бухгалтерам важнее указать конкретные счета и версии 1С, с которыми они работали.
Эксперты сервиса «Готовое резюме» составляют резюме без ошибок и учитывают особенности профессий. Час собеседования — и новый документ окажется у вас на почте и в личном кабинете, готовый к поиску идеальной работы.
Редактировать мое резюмеХочу готовое резюме!Топ 10 ошибок при составлении резюме на hh.ru и не только | про ошибки при составлении резюме, советы и рекомендации по составлению резюме | чек-лист для резюме
Про ошибки при составлении резюме
Рекрутеры не враги соискателям — в условиях дефицита времени на обработку больших массивов информации они оптимизируют свою работу, избегая тратить время на поиск и изучение резюме с непонятным и нелогичным содержанием.
Невозможно понравиться всем — это никому еще не удавалось, но сделать содержание резюме удобным для восприятия — несложная задача.
Ниже вы найдете перечень самых распространенных, на мой взгляд, ошибок при составлении резюме и рекомендации по их исправлению.
Надеюсь, мой Топ 10 поможет вам найти работу мечты.
Ошибка 1. Использование в качестве наименования желаемой должности названий несуществующих вакантных должностей
Если вас интересует вакансия «Менеджер по логистике», а в поле «Желаемая должность» вы пишете «Специалист по мультимодальным перевозкам», вы сами минимизируете ваши шансы получить желаемую работу, ведь самое распространенное название должности с таким содержанием работы — «менеджер по логистике».
И резюме работодатель в первую очередь будет рассматривать «менеджеров по логистике», а не «Специалистов…».
И при поиске резюме в базе работного сайта в поисковой выдаче по запросу «менеджер по логистике» сайт не либо покажет рекрутеру резюме «Специалист …», либо покажет в конце поисковой выдачи.
На месте рекрутера вы бы и сами не рассматривали или рассматривали в последнюю очередь резюме, наименование желаемой должности в котором не соответствует названию вакансии, особенно, если есть альтернативы в виде резюме с «правильными» названиями.
Совет 1
Не ленитесь до размещения резюме просмотреть интересующие вас вакансии и выберите для своих резюме наиболее популярные у работодателей наименования желаемых должностей.
Ошибка 2. Отсутствие в тексте резюме используемых в описаниях вакансий слов и словосочетаний
Рекрутер просматривает резюме с той же скоростью, с какой рядовой пользователь интернет просматривает вкладки браузера, открытые по результатам поисковой выдачи — на поиск нужной информации на вкладке тратится 6-20 секунд.
«Зацепить» может только соответствие текста резюме описанию вакансии — наличие в тексте резюме используемых в описаниях вакансий слов, терминов, словосочетаний.
Если их в резюме нет, поверьте, никто не будет читать про ваши профессиональные достижения, как бы цветисто они ни были описаны.
Хотите попасть на интервью — соответствуйте «поисковым запросам» работодателя.
О канонах составления резюме, подразумевающих использование глаголов действия и оцифровку достижений впечатляющими цифрами, забудьте — они были актуальны, когда резюме печатали на бумаге — сейчас его не читают, а просматривают в поиске соответствующих запросу ключевых слов.
Совет 2
Не ленитесь изучать описания вакансий, выбирайте из них распространенные слова, словосочетания, термины, используйте их в тексте резюме.
По технологии Search Engine Optimization (SEO) обрабатывается большинство контента, размещаемого в интернет, о том, как ее применять при поиске работы, я ранее писал в статье SEO оптимизация резюме для людей и роботов.
Ошибка 3. Неинформативный текст, большой объем резюме
Повторю то, о чем писал выше — на резюме рекрутер тратит 6-20 секунд.
Если вы готовы потратить 8 минут просмотр ролика, демонстрирующего работу рекрутера с резюме, посмотрите его на Youtube.
Ролик, сразу предупреждаю, нудный, но для понимания процесса вполне пригоден.
Если 6-20 секунд не хватит для того чтобы найти соответствующую поисковому запросу информацию, то в первую очередь винить в этом можно соискателя.
Поиск нужной информации в гуще таких неинформативных штампов, как «руководил деятельностью подчиненных» или «вел деловые переговоры» быстро утомляет, и, извините, раздражает рекрутера.
Поэтому резюме может быть закрыто, если на проверку его соответствия запросам работодателя нужно больше времени, чем готов потратить специалист по подбору персонала.
И не нужно обижаться на работодателей — пользователи интернета так же закрывают страницы, если у них не хватает терпения найти нужную информацию в объемном и неудобном для чтения тексте.
Совет 3
Сокращайте описание опыта работы на одном месте до 7-9 строк, неинформативные фразы безжалостно удаляйте.
Для того чтобы понять как рекрутер видит ваше резюме, почитайте статью Как рекрутеры рассматривают резюме в HH/Superjob — 7 скриншотов от HR-ПРАКТИКА.
Ошибка 4. Лишняя информация в резюме
Семейное положение, хобби, количество и возраст детей, религиозные и политические взгляды, воспевание соискателем своих выдающихся качеств не имеют отношения к соответствию опыта, знаний и навыков описанию вакансии, но могут быть неоднозначно и субъективно восприняты работодателем.
Подробнее о рисках размещения в резюме лишней информации читайте в статье про 40 причин, по которым соискателей не приглашают на интервью.
Кроме того, лишняя информация в резюме отвлекает работодателя от соответствующего его запросам текста.
Совет 4
Удалите из резюме информацию, не имеющую отношения к работе.
Если работодателю действительно важно знать, увлекаетесь ли вы дайвингом, дождитесь вопроса об этом на интервью.
Ошибка 5. Список разнородных/разноуровневых должностей в названии резюме
Резюме соискателя с названием искомой должности «Директор по продажам, менеджер по продажам», отправленное в качестве отклика на вакансию «менеджер по продажам», вряд ли будет рассматриваться — скорее всего, оно было отправлено по ошибке, ведь «директор» по определению не должен интересоваться вакансией «менеджер».
И резюме «Бухгалтер, специалист по закупкам» тоже замучает рекрутера сомнениями в истинных намерениях соискателя.
А вот резюме «Секретарь, помощник руководителя» подозрений, скорее всего, не вызовет — перечисление должностей с близким содержанием работы вполне логично.
Совет 5
Используйте несовместимые наименования желаемых должностей в отдельных резюме, еще лучше — публикуйте резюме с такими названиями в разных аккаунтах.
Ошибка 6. Несоответствие названия резюме опыту работы
Если резюме называется «коммерческий директор», но в опыте работы отсутствует опыт работы на аналогичной или близкой по содержанию работы должности, рекрутер не сможет убедить себя необходимости пригласить этого соискателя на интервью, ведь ничем, кроме желания занять искомую должность, амбиции соискателя не подкреплены.
Любая другая кандидатура с минимальным опытом на близкой по содержанию работы должности будет по определению предпочтительной.
Совет 6
Желание ставить перед собой амбициозные цели достойно уважения, но будьте реалистами — трезво оценивайте конкуренцию и ожидания работодателей на рынке труда.
Не стоит делать ставку на вакансии, шансы занять которые невелики — претендуйте также на вакансии, в конкурсе на занятие которых вы конкурентоспособны — лучше получать предложения и выбирать лучшее, чем тщетно ждать «работу мечты».
Ошибка 7. Несоответствие зарплатных ожиданий реалиям рынка труда
«Завышение» стоимости своей квалификации, конечно, хуже, чем занижение.
Если оказаться «не в рынке», предложений можно не дождаться.
Совет 7
Не ленитесь изучать рынок труда, сравнивайте свою квалификацию с квалификацией других соискателей, рассчитывайте свою стоимость на рынке труда.
О том, как это делать, я писал в статьях Как оценивать рыночную стоимость сотрудника или соискателя и Нестандартный кейс про оценку стоимости соискателя.
Ошибка 8. Ошибки в тексте резюме
Речевые, орфографические, синтаксические, стилистические.
Резюме — это реклама, вряд ли стоит рекламировать свою невнимательность и/или безграмотность.
Совет 8
Проверяйте и перепроверяйте текст, пользуйтесь проверкой орфографии в Word, просите вычитать резюме тех, кто силен в русском языке.
Ошибка 9. Неуказание профиля деятельности предыдущих работодателей
Если профильный опыт важен, стоит указать профиль деятельности бывших работодателей в начале описания опыта работы.
Не укажете — уменьшите свои шансы занять желаемую вакантную должность.
Совет 9
Укажите профиль деятельности предприятия сразу после наименования работодателя.
Ошибка 10. Неудачный выбор фото для резюме
Выбрать подходящее фото несложно.
Посмотрите на фото в резюме ваших конкурентов по рынку труда и подумайте о том, каким должно фото соискателя, который в представлении работодателя будет идеальным кандидатом на занятие вакантной должности, на которую вы претендуете.
Вот таким и должно быть ваше фото.
Совет 10
Подберите соответствующее резюме фото из личного архива или обратитесь к профессиональному фотографу.
И не злоупотребляйте Фотошопом.
Подробнее об этом в материале Как выбрать фото для резюме — рекомендации и кейсы от HR-ПРАКТИКА.
И еще два совета для пользователей HH.ru
Важно: HH.ru показывает наличие у соискателя нескольких резюме, если они размещены в одном аккаунте.
Совет 1
Разнородные/разноуровневые резюме размещайте в разных аккаунтах, HH.ru не запрещает заводить несколько аккаунтов.
Важно: «закрытие» резюме от своего текущего работодателя вовсе не означает, что он не сможет увидеть ваше резюме, посетив сайт в режиме неоплаченного доступа и «опознав» вас по содержанию резюме.
Совет 1
Если хотите исключить риск того, что ваш работодатель обнаружит, что вы ищете другое место работы, делайте резюме доступным только для тех компаний, на вакансии которых вы откликнулись.
В заключение
Буду рад, если коллеги расширят мой список ошибок и поделятся своими советами.
Если какие-то из рекомендаций выше показались вам лишком сложными для того, чтобы следовать им на практике, вам стоит изучить несколько статей с кейсами, в которых пошагово описаны примеры устранения ошибок в резюме.
Вот они — Кейс про составление резюме с HR-ПРАКТИКА, Как правильно составить резюме и 7 кейсов с рекомендациями по коррекции резюме от HR-ПРАКТИКА.
Желаю успехов в поиске работы и работников!
Автор: Денис Карандашев
Перейти в Каталог статей
Перейти в Каталог программ обучения
Перейти в Каталог консультационных услуг
Подписывайтесь и получайте анонсы свежих статей
Поделитесь ссылкой на статью с друзьями и коллегами
hh.ru — «Удаленная работа на hh.ru Как составить резюме, написать сопроводительное письмо, избежать мошенников и наконец-то найти работу на HeadHunter? Личный опыт, примеры и лайфхаки.»
Время от времени приходится искать новую работу и за пять лет я пришла к выводу, что большинство вакансий крупных фирм можно найти на HeadHunter, поэтому нет смысла мониторить несколько сайтов одновременно. Ведь чаще всего оказывается, что такое же предложение размещено на HeadHunter.
Как оформить резюме на HeadHunter
Сначала заполняются персональные данные, здесь для меня основной сложностью стал выбор фотографии, так как я не люблю фотографироваться, а ставить фото как в паспорте — слишком неприглядно, да и после опроса знакомых многие признались, что она от себя отталкивает. В итоге пришлось ставить фото, где я выгляжу гораздо моложе, но хотя бы смотрю в кадр и вызываю положительные эмоции. Никогда не возникало мысли, что можно обойтись без фотографии, ведь это дополнительный способ выделиться среди остальных.
У меня подтвержден телефон, для восстановления доступа и большего доверия. Я ищу удаленную работу в другой стране, поэтому предпочитаемый способ связи — электронная почта.
Я ищу удаленную работу в другой стране, поэтому предпочитаемый способ связи — электронная почта.
Забегая вперед расскажу об одной удобной функции «Видимость резюме». Чтобы избежать пустых предложений и постоянных звонков, я скрываю резюме из поиска. Возможно, так я пропущу заманчивую должность, но чаще всего это будет неинтересная и скучная работа. Лучше самому выбирать, куда отправляешь отклик, чтобы не тратить время.
Параметр «Анонимное резюме» позволяет скрыть контактные данные, эту настройку я в полной мере оценила только сейчас. Тратиться на роуминг нет смысла, ведь вопросы можно обсудить и почте, а интервью пройти по скайпу.
 Как заполнить поле опыт работы и графу обязанности
Как заполнить поле опыт работы и графу обязанностиДля себя я решила, что нет смысла указывать опыт работы меньше нескольких месяцев, так как придется объяснять, почему пришлось уволиться. Также я считаю, что это плохо влияет на общее впечатление от резюме. На первых порах в этом блоке мне нечего было написать, тогда я просто указала, что проходила практику от университета.
У меня четыре разных резюме, в каждом указан опыт работы, который больше подходит по профилю. Исключение делаю только для одной записи о трудоустройстве, которая присутствует во всех вариантах, потому что там большой опыт работы. Также в резюме отмечено, почему я ушла из крупной компании спустя четыре месяца, во избежание лишних вопросов.Лайфхак, как заполнить графу обязанности.
Я делаю так — нахожу несколько аналогичных вакансий и выбираю самое лучшее, пару раз даже копировала информацию из вакансий предыдущего места работы А чтобы не писать лишнего, указываю именно те обязанности, которые будут востребованы на новой работе.
Еще одна нужная для заполнения графа — ключевые навыки. Здесь указываются подходящие для будущей работы умения. После ввода нескольких слов система дает подсказку, подбирая похожие навыки. Я стараюсь не перегружать эту строчку информацией и пишу не больше 10 навыков.
Никогда не заполняла пункт «Обо мне», где предлагается рассказать о хобби или личных качествах, считаю, что остальной информации вполне достаточно для начала. Гораздо больше значения имеет сопроводительное письмо, которое повышает вероятность прочтения резюме. Остальные пункты стандартные и не вызывают затруднений.
Еще есть возможность указать несколько дополнительных параметров, возможно, для крутых специалистов они имеют значение, а мне пока нечего написать.
В целом, мне очень нравится форма для резюме на сайте, очень понятно и удобно, а также можно настроить параметр «Видимость», который облегчают жизнь после размещения резюме.
Как составить сопроводительное письмо
В большинстве случаев при отклике на вакансию я отправляю сопроводительное письмо, чтобы показать свою заинтересованность и привлечь внимание к резюме. Если мне не нравится вакансия по описанию, но привлекает компания или возможности, то я оставляю отклик без письма.
Перед тем, как составлять сопроводительное письмо, я прочитала много советов из разных источников. Стандартный шаблон выглядит так:
Написано просто, но когда я села писать письмо по этой схеме, то впала в ступор. Ничего толкового не приходит в голову, а если расписать все указанные пункты, выходит слишком громоздко и уныло. Поэтому я решила искать нестандартных советов и примеров, которые в итоге помогли составить хорошее сопроводительное письмо. И они сводятся к двум рекомендациям, которые помогают по-другому взглянуть на структуру письма:- Ответьте на два вопроса — почему мы, почему вы.
- Отвечайте так, чтобы полностью закрыть потребность в вакансии. (если в вакансии написано, что нужен секретарь с высокой скоростью печати, пишите об этом, а не о знаниях языка).
 (если в вакансии написано, что нужен секретарь с высокой скоростью печати, пишите об этом, а не о знаниях языка).
(если в вакансии написано, что нужен секретарь с высокой скоростью печати, пишите об этом, а не о знаниях языка).Пример сопроводительного письма, благодаря которому мое резюме просматривают в 90% случаев.
Как искать работу с помощью HeadHunterНа этапе настройки фильтров все легко, указал желаемые параметры и вперед. Единственный минус — нельзя указать несколько параметров сразу. Например, для опыта работы никак не выбрать сразу два пункта «нет опыта» и «от 1 года до 3 лет». Дело в том, что в последней категории много вакансий, в описании которых указано, что опыт работы приветствуется, но необязателен. И приходится искать по очереди, что неудобно.
Дальше идет отбор вакансий, еще одна удобная функция — скрыть из поиска все вакансии выбранной компании или только одну. В этом списке у меня салоны сотовых сетей, банки, такси и вакансии авторов работ, так как их тьма (на каждый город по каждой специальности).
Особенности поиска удаленной работы на HeadHunter
Здесь все не так просто, как кажется. Потому что многие вакансии, которые не подразумевают удаленную работу, в глазах работодателя почему-то приобретают такой смысл. И появляются в выдаче таксисты, швеи, повара и прочее. Для меня удаленная работа — это возможность работать из любой точки мира. Поэтому мне здорово помогает оператор not, забиваю в поисковую строку список исключающий слово и спокойно смотрю новые вакансии.
Вот пример списка, который я копирую в поисковую строку, чтобы сразу отсеять лишнее: not звонки not телефон not колл-центр not программист not бухгалтер not юрист not водитель not входящие not переводчик not программист.
На какие вакансии не стоит откликаться
Я с сомнением отношусь к вакансиям по набору текста, бесплатному обучению трейдеров или другой, подозрительно легкой работе с высокой оплатой, особенно, если отвечают на запрос быстро и просят зарегистрироваться на каком-нибудь специальном сервисе. В интернете часто находятся негативные отзывы о работе такого сервиса или после проверки сайта, оказывается, что домену всего месяц, а компания заявляет о 10 летней работе на рынке труда. Еще один звоночек — у компании нет вакансий по другим направлениям.
Подводя итоги — мне нравится этот сайт, с помощью него я не раз находила работу, интуитивный интерфейс, бесплатное размещение. Конечно, на сайте предусмотрено много платных сервисов и услуг, но я ими никогда не пользовалась и не планирую, поэтому мне нечего о них рассказать. И последний замеченный минус — нет кнопки пожаловаться на вакансию или на работодателя. Иногда хочется оставить жалобу на сомнительную вакансию, но для этого, видимо, нужно написать в службу поддержки.
_______________________________________________________________________________
Другие отзывы по этой теме:
Про удаленную работу онлайн-консультантом
Получаю зарплату на карту Альфа-банка, снимаю деньги без комиссии заграницей
Предвзятый и субъективный взгляд на резюме разработчика
Привет! Недавно мои коллеги из hh.ru провели вебинар, где разбирали резюме разработчика. Я решил поделиться своей точкой зрения на этот счет.
В этой статье мы поговорим о том, как разработчики создают резюме. Я расскажу о том, что меня, как интервьюера, мотивировало и воодушевляло перед встречей с кандидатом, а что наоборот. Разумеется, предвзятость в начале собеседования — это плохо, но все мы люди, и не можем полностью абстрагироваться от входных условий.
Дисклеймер: всё, что я буду описывать в этой статье — субъективно и базируется на опыте и эмоциях, которые я испытываю, когда читаю резюме, собеседую разработчиков etc.
Если true story читать не хочется, а хочется сразу tl;dr (в виде “Итого”) или how-to (в виде гайда), то можно промотать вниз на два последних блока.
Оффтоп: зачем мне тратить время на эту статью?
Давным-давно, еще студентом, я сидел перед компом и пытался воткнуть в это самое «нужно создать резюме, чтобы рассказать, какой я хороший и замечательный, и почему меня нужно взять без опыта». Вооружился гуглом и искал материалы формата «как создать резюме, в которое влюбится работодатель». Получалось примерно следующее: «Стрессоустойчивый кандидат познакомится с привлекательным работодателем. Хочу опыт и кусок хлеба, могу в потуги писать на C#, вот даже в универе такую штукенцию забахал!».
Резюме образца 2013 года
Почему я тогда указал Perl — я не знаю, не спрашивайте. Я написал на нем 2 строчки для лабораторки в ВУЗе и больше никогда не открывал.
Дальше было так: я сходил на собеседование в Яндекс на позицию фронтенд-разработчика в команду переводов. Там я красиво провалился. Я не знал вообще ничего. Сверстать пару блоков и немного покодить на JS я умел, конечно. Но в теоретической части я был где-то в районе -1.
Из того собеседования я вынес 3 мысли:
- Я хочу быть фронтендером, потому что они крутые и РИСУЮТ САЙТЫ. Что ж, мне было 19 лет, поэтому такие суждения простительны.
- Я хочу работать в яндексе. Там очень умные ребята, которые очень крутые. Позднее в Яндекс я всё-таки прошел и даже получил офер, но остался в hh.ru.
- Я провалил вопрос про выделение текста в textarea. Через несколько месяцев этот вопрос снова стал меня мучать, и я написал свою первую полноценную статью про JS на эту тему.
Дальше была Школа Программистов hh.ru, где из меня и сделали человека разработчика.
Это была лирика. Далее по теме
За время работы в hh.ru я участвовал в огромном количестве собеседований. Сначала как подмастерье, а потом уже как ведущий этого праздника жизни. Мне всегда казалось, что собеседовать нужно совсем по-другому, нужно лучше. И в какой-то момент я стал много времени уделять резюме кандидата.
Я взял за правило, что к моменту собеседования резюме должно быть прочитано полностью, а во время интервью может использоваться только как набор закладок и шпаргалки для разговора.
Зачастую резюме меня удивляли. После их прочтения у меня складывалось ощущение, что этот человек точно нам подойдет. Они не обязательно были уровня «работал в Netflix 100 лет, но решил вернуться в Россию». Было нечто другое, что “цепляло”. Так я понял, что у резюме несколько задач:
- Успешно пройти ПВО в виде HR-менеджера
- “Зацепить” интервьюера, чтобы собеседование не скатилось в стандартное обсуждение 1000 и 1 вопроса, необходимых на интервью
Однако далеко не всегда интервьюер будет читать ваше резюме перед собеседованием. Я даже провел небольшой «эксперимент»: сходил в N компаний на собеседования в 2018-2019 годах. Сами собеседования условно можно было разделить на два типа:
- Интервьюер просто делает свою работу, ты ему вообще до лампочки. Здесь все понятно.
- Интервьюер ищет себе коллегу. Здесь и начинается диалог, обсуждаются технические моменты, а уходишь с ощущением, что всё было отлично. Офферов было немало, но мне нравится работать в hh.
Опираясь на опыт проведенных мной собеседований и большого количества рассмотренных резюме я и хочу построить дальнейший рассказ.
А чтобы всё было по-честному: где-то в конце 2019 я угробил 4 выходных дня, чтобы выстрадать свое резюме.
Успешно пройти иммунную систему в виде HR
На мой взгляд, базовый инструментарий HR-менеджера включает в себя:
- Договоренности с техническим департаментом, на что обращать внимание. Часто можно найти в wiki компании или просто спросить об этом HR. Например: «если в опыте кандидата верстка промиков — это не наш профиль» или «если работал над почтовыми сервисами — это нам нужно, зовем».
- Личный опыт. Набор личных эвристик основанных на предыдущем опыте: «ребята из этой компании сильные, другие разработчики о них хорошо отзываются», либо «кандидат меняет работы каждые полгода, стоит почитать его резюме внимательнее и желательно между строк».
- Keywords: React, Redux, Spring, Java, 15 лет опыта, Node.js
- Ачивки. «Съел пирог и не растолстел», «Моя фича набрала +100% к пользователям»
Зацепить интервьюера
Резюме — это краткое сочинение на тему «почему я хороший». Цель разработчиков — рассказать о тех вещах, которые могут зацепить.
Например:
- Делал клевый проект или отвечал за весь слой? Напиши про это.
- Занимался проработкой задач вместе с бизнес-заказчиками? Пара слов про это даст понять интервьюверу, что вы работаете не только на уровне «таска пришла-таска ушла»
- Есть что-то «около-работы»? Отлично, черкани пару строк. Нет такого? Да не проблема, life-work balance тоже ок.
Помимо «почему я хороший», резюме — это еще и набор тем для общения:
- Занимался архитектурой проекта? Отлично, можем похоливарить!
- Занимался перформансом? Круто, интервьюер увидит, что кандидат в этом сечет. А еще это повод начать не со стандартного «картинки нужно сжимать, а js тяжелее img», а с каких-то практических примеров из опыта.
Опыт работы. Препарируем пример
Крайне разочаровывают резюме уровня «работал работу на работе». Здесь отсутствует человек, есть просто функция. Приведу пример:
Компания: "Соло на трекпаде",
Frontend-разработчик, Июль 2018 — по наст. бремя.
Участие в разработке продуктаНичего не понятно. Есть портал. Он внутренний? Внешний? А сайт? Приложение? Чат? Какова в этом роль кандидата? Возможно было что-то, что в проекте нравилось? И наоборот? Миллиард вопросов!
Стоит чуть-чуть пройтись шпаклевкой, и такой пример становится полезнее:
Компания: "Соло на трекпаде",
Frontend-разработчик, Июль 2018 — по наст. бремя.
Участие в разработке продукта для общения с клиентами через чаты техподдержки.
- Интеграция с телеграм, WhatsApp и вайбер
- Аналитика данных
- АБДля меня, как для интервьюера, становится понятно, что разработчик выделил для себя. Отличный повод поговорить о том, как проводились АБ тесты и собирались метрики. Необходимо понять, насколько разработчик — разработчик, и насколько мы на одной волне.
Для достижения лучшего эффекта кандидат может расставить акценты на том, что его драйвило:
Компания: "Соло на трекпаде",
Frontend-разработчик, Июль 2018 — по наст. бремя.
Участие в разработке продукта для общения с клиентами через чаты техподдержки.
- Интеграция с телеграм — написали свой клиент через SDK.
Это позволило увеличить конверсию пользователей на 100 человек в день
- Аналитика данных, АБ — организовал инфраструктуру для связи "Компоненты - эксперименты"Кажется, что описано то же самое, что и в предыдущем примере, но:
- Интервьюер уже видит, что кандидат не дядя с улицы, а занимается вопросами улучшения и оптимизации. Чтобы проверить свою гипотезу, я завожу разговор о той самой инфраструктуре. Выясняется, что информация о том, от какого эксперимента зависит компонент, находится в static-поле. Бабель-плагин достает такие поля и формирует JSON страница ⇒ эксперименты.
- Упоминание достижений, даже локальных, говорит о том, что разработчику «не все равно». Не все равно, что там бизнес творит. Не все равно на фидбек сервиса. 100 очков гриффиндору.
Появляется логичный вопрос: А если так?
Компания: "Соло на трекпаде",
Frontend-разработчик, Июль 2018 — по наст. бремя.
Участие в разработке продукта для общения с клиентами через чаты техподдержки.
Используемый стек: React, Redux, Кин-дза-дза, Node.js, FusRoDuh v3.Это не даст вам преимущества. Упомянуть о стеке хорошо. Но превращение всего опыта работы в стек «я кодер, кодерю на стеке» не говорит о том, что разработчик умеет решать задачи.
Можно выбрать какой-нибудь срединный путь:
Компания: "Соло на трекпаде",
Frontend-разработчик, Июль 2018 — по наст. бремя.
Участие в разработке продукта для общения с клиентами через чаты техподдержки.
- Интеграция с телеграм — написали свой клиент через Node.js SDK.
Это позволило увеличить конверсию пользователей на 100 человек в день
- Аналитика данных, АБ — организовал инфраструктуру для связи "React компоненты - эксперименты"
- Организация бизнес слоя на Redux + Saga, иницировал переход с thunk на SagaЗдесь в каждый пункт мы добавили немного контекста: вот тут я трогал ноду, здесь реакт, а еще и полностью техническим вопросом занимался! Танки на саги перетаскивал.
В итоге: Мы увеличили наш текст на 3 пункта, но сделали его с одной стороны соответствующим для HR менеджера, который и keywords увидит, и человека рассмотрит. А с другой стороны он хорош для разработчика — собеседующий будет понимать, что интересно разработчику (ну или чем его заставляли заниматься).
Гайд по заполнению резюме
Начнем со сложного: сколько писать? Когда я хочу написать коротенькую статью “буквально на 3 абзаца”, выходит вот это. С резюме так же. В какой-то момент мое резюме занимало 4 страницы А4.
Идеально, если получится впихнуть резюме в одну страницу. Что там можно написать?
Кто таков. Имя-фамилия-позиция-контакт. Фото и прочие прибамбасы — по желанию. Тут как с авто — обвес лишним не будет, но и без него едет.
Не оч:
Аноним Анонимов
[email protected]Уже лучше:
Мария Игнатова, Frontend-разработчик
Telegram: @definiteTelegramAccount
email: [email protected]Кайф:
Мария Игнатова, Frontend-разработчик
Telegram: @definiteTelegramAccount
email: [email protected]
GitHub, LinkedIn links, etc.Чем славен/tl;dr/эбаут. Часто в «о себе» либо ничего не пишут, либо пишут о стрессоустойчивости. А я бы порекомендовал вынести туда самую мякотку: «Lead developer, водил команду из 6 человек в данжи. Отвечал за направления чат-ботов. Автор статей на Хабре.» Сюда же можно засунуть «пожелания». Например: «хочу работать в игорном бизнесе, энтерпрайз надоел, галеры ван-лав.» В этом случае эбаут можно разделить на две части. Мякотка вверху, пожелания внизу.
Можно лучше:
Frontend engineerУже лучше:
Frontend engineer.
Нравится BDD, прорабатывать архитектуру,
заниматься глобальными вопросами в проекте.
Хочется совмещать решение сложных технических задач и продуктовую разработку.
В продуктовой разработке люблю заниматься вопросами метрик, организации процессов.Другой вариант:
Frontend engineer. Автор технических статей. Спикер на РИТ++, FrontendConf.
Сосредоточен на решении продуктовых задач.И третий вариант:
Frontend engineer
Есть опыт разработки продуктов с нуля и развития комплексных проектов.
Хотелось бы попробовать себя не только во фронтенде,
но и в форточку выкидывать свои навыки не хочу.
Не заинтересован в игорном бизнесе.Опыт работы. Об этом выше уже было (сразу перед этой главной), поэтому идем дальше
Около работы. Статьи, выступления, опен-сорс, пет-проджекты, волонтерства, я у мамы молодец — об этом забывать не стоит. Около-работа ощутимо влияет на принятие решений. Однажды я собеседовал разработчика, который написал, что сделал промисы, чей колбек для исполнения выполняется в отдельном воркере. Это дало повод пообщаться о том, как это работает. Этот разработчик вполне успешно работает у нас уже несколько лет. Как говорится, “все, что вы скажете, может быть использовано против вас”. Важна каждая деталь. Если есть чувство, что об этом говорить не хочется или нечего, то блок пропускается и никто ничего не заподозрит.
Нормально:
Дает больше контекста:
Автор небольших опенсорс-проектов,
которые выделили во время работы над рабочими задачами:
например https://github.com/hhru/babel-plugin-react-displayname
Спикер на конференциях. Например РИТ++ (ссылка на доклад)Другой вариант:
Прохожу курсы на egghead, coursera,
Нравится посещать конференции, делиться знаниями
Провожу митапы внутри компании.Keyskills. Не будем усложнять серчерам задачу. Лучше выделить блок, который можно оформить или просто как поток сознания, или его же, но с временными рамками:
Поток сознания:
TypeScript, React, Redux, Svelte, Babel, Node.js, Koa, Express, JS, CSS, HTMLПоток сознания с временными рамками:
React, Redux, TS — 3+ года
JS, CSS, HTML — 6+ лет
Node.js — 1 годЯ предпочитаю первый вариант. Он компактнее и не засоряется лишней информацией.
Общая разлиновка. Мне нравится такая:
Да, я бессовестно содрал со своего же резюме, но зачем мне рекламировать то, что мне не нравится?
Однако можно не заморачиваться, и воспользоваться готовым сервисом, который поможет составить резюме, а в лучшем случае и работу найдет. Я в разное время заполнял резюме на этих сервисах (ln не в счет, он — соцсетка):
- https://www.jobhero.com/ — много разных шаблонов, приятный интерфейс, здесь я и взял вдохновение для своего резюме, владельцы забанили российские IP.
- https://hh.ru/ — ну, вы поняли
- https://headz.io/ — что-то вроде российского аналога hired/honeypot со своим видением вопроса. Удобно: заполнить резюме можно быстро, легко и без смс. Неудобно: keyskills и «в какой хочешь работать компании» немного занудные
- https://www.honeypot.io/ — они крайне назойливо хотят созвониться, так как на основе твоего резюме будут подбирать тебе работу. В остальном — зайчики
- https://hired.com/ — Все ок, но у меня не получалось отправить резюме на ревью
- https://indeed.com/ — очень порадовал удобным интерфейсом для заполнения резюме
(список составлен в рандом порядке и смысла не несет. Профиль на хабр-карьере не трогал с 2018 года, судить не могу)
Итого
Далее пойдут сухие советы без кринжового веселья.
Вещи которые нам нужны в резюме:
- Контакты.
- Что ты из себя представляешь, что нравится в разработке: достижения на проекте, пусть это будут и совсем локальные, возможно то, что нам греет сердце. Обычно включается в опыт работы
- Контекст проекта — чем занимался, в идеале, если есть опорные пункты. Опорные пункты вида: «сделал фичу календарь; улучшил перфоманс» и т.д. Такие штуки выполняют 2 задачи: показывают со стороны специалиста и позволяют задать тон будущему собеседованию (но зависит от опыта собеседующего). Тоже включено в опыт работы
- keywords — хоть мне это и не нравится, но на те самые «ключевые навыки» обращают внимание. Способов их вставить всего два. Первый — нативно во время рассказа про контекст проекта/достижения. Второй — отдельный пункт «Ключевые навыки». Тем самым мы упрощаем время скрининга нашего резюме и повышаем шансы не быть отложенными по принципу «ну мы тут Angular ищем, а кандидат про это не упомянул»
Вещи которые включаются по желанию:
- О себе. Нравится проводить митапы? Участвовать в них? Написал статью на Хабре? Работаешь с пет-проджектами? Коммитишь в опен-сорс? Ничего из перечисленного, но было что-то интересное на работе, что можно перенести из описания проекта в блок о себе? Снова нет? Можно опустить его, отрицательно это не скажется.
- Пожелания. Был неприятный опыт с командой? Есть определенные убеждения? Это можно указать в конце резюме.
И в заключение стоит сказать — относитесь к резюме не как к официальной бумаге, а как к вашему «элеватор питчу«.
Образец сопроводительного письма к резюме 2021
Правила составления
Нет конкретных установленных требований к этому документу, но существуют определенные пункты, которые традиционно присутствуют в сопроводительном письме. Предлагаем вам несколько примеров, как написать сопроводительное письмо к резюме. Всегда в нем стоит указывать:
- На какую вакансию вы претендуете.
- Чем вас заинтересовала вакансия.
- Почему вы хотите работать у этого работодателя.
- Чем вы можете быть интересны работодателю (краткое описание опыта, профессиональных навыков и компетенций).
- Ожидаемый уровень дохода (если он не указан в вакансии).
Ну и, конечно, помимо содержательной части, то есть непосредственно того, что писать в сопроводительном письме к резюме, не стоит забывать и о классических правилах деловой переписки. Текст начните вежливым приветствием и обращением (если вы знаете конкретные имена), а закончите вашим именем и контактными данными («С уважением, Иванов Иван, номер телефона +7 (ХХХ) ХХХ-ХХ-ХХ»).
Если резюме отправляется по электронной почте, то сопроводительное письмо размещают в теле письма. Если резюме отправляется через сайты по поиску работы (например, hh.ru или superjob.ru), то для текста сообщения существует выделенное поле. Также на сайтах по поиску работы есть возможность настройки текста сопроводительного письма.
Примеры
Хотя общие правила составления документа понятны, конкретные образцы всегда удобны, потому что их легко адаптировать под себя.
Пример краткого сообщения:
Добрый день, Мария Ивановна. В ответ на Вашу вакансию «электрик», размещенную в газете «Работа», высылаю свое резюме. Буду признателен, если Вы не оставите его без внимания.
С уважением, Иванов Иван. Телефон для связи +7-123-456-78-90.
Развернутый пример сопроводительного письма к резюме, hh.ru (форма на сайте):
Даже если вы планируете составить развернутое обращение, старайтесь не перечислять все пункты резюме. Пишите об опыте кратко, но так, чтобы было понятно, что конкретно вы делали. В качестве примера — вариант сопроводительного письма к резюме на вакансию юриста.
Здравствуйте!
Ваша компания опубликовала вакансию на замещение должности юриста в газете «Работа». Мой юридический стаж составляет 5 лет, и мне бы хотелось предложить Вам свою кандидатуру. Я окончил юридический факультет Института юридических наук г. Санкт-Петербурга, хорошо разбираюсь в отраслях гражданского и трудового законодательства. Работал юрисконсультом в ООО «Весна» (г. Санкт-Петербург), ООО «Пион» (г. Санкт-Петербург), вел делопроизводство, участвовал в работе по подготовке и заключению хозяйственных, трудовых и коллективных договоров, консультировал работников организаций по организационно-правовым и иным юридическим вопросам.
Хорошо владею современными офисными программами и правовыми системами. Среди моих качеств — доброжелательность, ответственность, стремление вникнуть в суть проблемы, стрессоустойчивость.
Надеюсь, Вас заинтересовало мое резюме. С удовольствием приму предложение встретиться и рассказать больше об опыте работы и возможном потенциале. Вы можете связаться со мной по телефону +7-123-456-78-90 либо электронной почте [email protected]
С уважением, Иванов Иван
Другой пример — сопроводительное письмо к резюме администратора:
Добрый день!
Меня заинтересовала вакансия администратора в вашей компании, опубликованная на сайте rab.ru. Я внимательно изучил указанные требования и уверен, что полностью соответствую им.
Имею высшее образование в туристической сфере, что соответствует профилю вашей компании, и хочу предложить свою кандидатуру на ваше рассмотрение. Работал администратором в отеле «Краснодар» (г. Краснодар), гостинице «Сочи» (г. Сочи), в туристическом агентстве «Путешествуйте с нами» (г. Анапа).
Среди моих полезных навыков — умение работать с документацией и клиентами. Вел клиентскую базу, занимался консультациями, проводил телефонные переговоры с клиентами. Владею всеми пакетными компьютерными программами. Мой стаж в сфере менеджмента составляет 7 лет.
Готов прийти на собеседование в указанное Вами время. Вы можете связаться со мной по телефону +7-123-456-78-90 либо электронной почте [email protected]
Надеюсь на сотрудничество.
С уважением, Иванов Иван
Хорошо, если у вас есть хоть какой-то опыт, всегда можно написать о нем. Но если его в конкретной сфере пока нет, сообщение об этом для отдела кадров еще полезнее. Вот пример сопроводительного письма к резюме для соискателя без опыта работы в конкретной сфере.
Добрый день!
Я ознакомился с предложенной Вашей компанией вакансией на замещение должности менеджера по продажам. У меня отсутствует требуемый опыт по этой специальности, но я обладаю целеустремленностью и готов изучать новое для меня направление. Мне очень хочется профессионально развиваться и повышать уровень знаний. Среди моих качеств — упорство, умение планировать, способность убеждать людей.
Буду признателен, если вы пригласите меня на собеседование. Мои контактные данные: телефон +7-123-456-78-90, электронная почта [email protected]
С уважением, Иванов Иван
Почему нужно иметь резюме не в формате hh.ru
Итак, резюме не в формате hh нужно как минимум в двух случаях:— Когда вы составляете резюме с нуля – чтобы избежать грамматических и пунктуационных ошибок.— И когда вы отправляете резюме работодателю напрямую – то есть ищете работу через знакомых, откликаетесь своим резюме на вакансии, размещенные на карьерных страницах интересных вам компаний, ищете вакансии в группах поиска работы в соцсетях.
Как показывает практика наших клиентов, если речь не идет о супервостребованных специальностях, таких как, например, разработчики, работа только в половине случаев находится на hh. В остальном это результат поиска по знакомым и группам в соцсетях.
Хорошо, скажете вы, если резюме должно быть не в формате hh, то в каком формате?
Выбирая свободный формат, соискатели тоже делают ряд ошибок. Например:
— Вставляют в резюме таблицы. Ряд компаний обрабатывает резюме в системах, автоматизирующих процесс подбора. Как правило, в них таблицы плывут и резюме выглядит крайне неэстетично, и это впечатление сразу переносится и на профессиональные качества кандидата. Поэтому рекомендуем убрать из резюме таблицы.
— Находят красивые шаблоны оформления резюме в интернете с диаграммами, рисунками, фоном и так далее. Как правило, и эти форматы плывут при автоматической обработке, а иногда появляются незаполненные фрагменты страницы, что очень раздражает подборщиков: им приходится вручную форматировать текст, заполняя пустые части листа, чтобы при печати не тратить лишнюю бумагу.
— Высылают резюме в формате pdf, что тоже серьезно портит подборщику жизнь. Как правило, рекрутеры перед пересылкой резюме нанимающему менеджеру вносят необходимые изменения в резюме: убирают ошибки и те моменты, которые не понравятся руководителю, добавляют важную информацию, которую они узнали в телефонном интервью. Но не все умеют вносить изменения в pdf и мучаются, переводя резюме в word, или так и не вносят необходимые им изменения.
Как вариант, можно использовать стандартные форматы оформления резюме, которые наряду с другими шаблонами предлагаются в Word.
Отчетность— Создание отчетов — База знаний HH Timing
В настоящее время доступно 3 типа отчетов в режиме HH Timing:
- Быстрый отчет
- Автоотчет
- Пользовательский отчет
Назначение быстрого отчета — скопировать дисплеи, открытые в данный момент в вашем проекте, в отчет HTML. Фильтры и конфигурация каждого дисплея в проекте будут учитываться и тиражироваться при создании отчета.
Кнопка доступа к быстрому отчету находится на вкладке «Отчеты», как показано ниже:
При нажатии кнопки «Быстрый отчет» каждый дисплей копируется в файл HTML и автоматически открывается в браузере пользователя по умолчанию. При необходимости этот файл можно сохранить для совместного использования в браузере.
В следующем примере макет дисплея в проекте включает:
- Главное табло
- График растущего среднего
- Табло для машины №1 (за графиком повышающихся средних)
- Табло лучшего сектора
Каждый из них будет скопирован в отчет HTML.
Примечание будут распечатаны только дисплеи, относящиеся к отчету, например, карты треков не будут добавлены в отчет.
Автоматические отчеты
предназначены для быстрого создания подробных отчетов стандартного формата с минимальным вмешательством пользователя. Чтобы создать новый профиль автоматического отчета, нажмите кнопку «Новый профиль» на вкладке «Отчеты».
Это даст пользователю возможность назвать профиль отчета. Здесь одновременно будут показаны все существующие профили отчетов в текущем проекте.
Обратите внимание, что после создания нового профиля его можно будет найти в дереве проекта слева вместе с другими существующими профилями отчетов в проекте.
После нажатия кнопки «ОК» новый профиль отчета откроется на титульной странице «Конфигурация отчета». Здесь можно ввести данные, которые будут отображаться на титульной странице, как показано ниже. Любые номера автомобилей, введенные в поле «Автомобиль (а)», будут суммированы в начале отчета в таблице с указанием водителей и цветом отчета.
Обратите внимание, что для включения титульной страницы в отчет необходимо установить флажок «Включить титульную страницу, но установить флажок».
Обратите внимание, что поле «Заголовок отчета» определяет имя файла, который будет сохранен.
Чтобы определить содержание отчета, выберите вкладку Автоотчет. Нажмите кнопку «Добавить главу» на панели управления на ленте отчета, чтобы добавить первую главу в отчет. Глава обычно используется для каждого сеанса, поэтому в одном отчете для каждого сеанса будет определена глава:
- Каждая бесплатная практика
- Квалификация
- Гонка
Когда создается новая глава, она выглядит так, как показано ниже.
Структура отчета
Структура отчета определяет тип шаблона для главы. В настоящее время доступны конструкции:
- Гонка на выносливость
- Сессия на выносливость на время
- Сводка по нескольким сеансам
Обратите внимание, что в сводке по нескольким сеансам можно выбрать несколько сеансов, тогда как в других структурах отчета можно выбрать только один сеанс.
Название главы
Определяет название главы.Например, FP1.
Выбор сеанса
Определяет, о каком сеансе будет сообщаться в этой главе
Выбор автомобиля
Определяет, какие классы будут включены в главу. Их можно расширить, чтобы выбрать / отменить выбор определенных автомобилей.
Focus Cars
Определяет любые автомобили, которые необходимо включить в более подробный анализ. Обычно это используется только для машин команды. Включены некоторые дополнительные сюжеты и выделены таблицы для автомобилей в фокусе.
Копировать выбранный автомобиль во все главы
Копирует выбор автомобилей из этой главы во все главы отчета. Это экономит время на выбор автомобиля, когда в отчет включается много глав.
Пример
В приведенном ниже примере в отчет включены 7 глав, по одной для:
- Сводка по всем сеансам
- FP1
- FP2
- FP3
- FP4
- квартал
- Гонка
Как показано, в главе «Сводка по всем сеансам» используется структура отчета «Сводка по нескольким сеансам» V2 , и выбраны все сеансы за выходные.В главе FP1 выбрана только сессия FP1. Все автомобили в классе Prototype были отобраны, и ни один Focus Cars не определен.
Чтобы создать отчет, нажмите кнопку «Создать автоматический отчет» на панели управления отчетом. Отчет откроется автоматически в форматах, определенных в конфигурации отчета (HTML, PDF или оба).
Они также будут сохранены локально на компьютере в следующем каталоге:
C: \ Users \ ИМЯ ПОЛЬЗОВАТЕЛЯ \ Documents \ HH Timing \ Reporting
Пользовательский отчет похож на Быстрый отчет в том, что он основан на дисплеях и данных, которые в настоящее время активны в проекте.Разница составляет:
- Титульный лист может быть определен так же, как Автоотчет
- Отчет сохраняется в том же каталоге, что и автоматические отчеты при создании
- Пользователь может контролировать порядок отображения каждого дисплея
- Отображение может быть удалено из отчета
- Формат сохранен и может быть повторно использован в будущем с другими сеансами
Находясь на вкладке «Пользовательский отчет», нажатие кнопки «Заполнить отчет» скопирует все дисплеи, которые в данный момент активны в проекте.Пример этого показан ниже:
На этом этапе порядок или каждый дисплей и то, что включено, могут быть изменены. Нажмите кнопку «Создать настраиваемый отчет» на панели управления лентой отчета, чтобы создать отчет. Отчет будет автоматически открыт в форматах, определенных в конфигурации отчета (HTML, PDF или оба).
Они также будут сохранены локально на компьютере в следующем каталоге:
C: \ Users \ ИМЯ ПОЛЬЗОВАТЕЛЯ \ Documents \ HH Timing \ Reporting
сводок в столбцах со значениями времени в чч: мм: сс
Отправлено 31 августа 2009 г. в 15:16
Я не программист на Java.У меня есть отчет, который должен вычислять список значений времени в сводках групп. У меня есть другие столбцы с различными числовыми типами данных, которые хорошо резюмируют. Однако эти значения времени не суммируются. Вместо этого я просто получаю последнее значение в группе, отображаемое в итоговом поле группы.
Использование JasperServer CE 3.5. Использование iReport 3.5.2. Данные поступают из приложения на базе Microsoft, которое принимает тип данных MS Double и форматирует его в формате чч: мм: сс. (MS использует двойное число, где целая часть числа = # дней, а десятичная часть = доля дня.)
Эти отформатированные данные не суммируются, и я предполагаю, что их невозможно вычислить. Я помню, что при построении отчета по умолчанию использовался тип данных Timestamp. Во всяком случае, в настоящее время он находится в поле отметки времени. Для ухмылки я тоже попробовал это как тип данных Time, но безуспешно.
Итак, возникает один вопрос: можно ли резюмировать тип данных Timestamp, и если да, то как?
Я попытался изменить запрос данных, чтобы создать поле с длинным значением: умножение значения поля с двойной точностью * 86400000 [число мс в день] и удаление значений справа от десятичного числа, и значение получится как Давно в отчете Джаспера.Я предположил, что Long — это то, что нам нужно, поскольку справка Java показывает, что конструктор Timestamp принимает значение Long (= количество миллисекунд). Опять же, я не программист на Java, поэтому я могу неправильно интерпретировать ситуацию …
Во всяком случае, мне не удалось получить выражение текстового поля в сводке группы, используя для компиляции класс java.sql.Timestamp. Я перепробовал столько разных вариаций темы, что не стоит выкладывать то, что я пробовал. Наиболее частые ошибки — это ошибки «не могут быть устранены».
Второй вопрос — как мне получить список раз для суммирования в столбце, а затем отобразить предпочтительно в формате чч: мм: сс?
Зарегистрирован: 2 апреля 2009 г. — 17:03
Последний визит: 12 лет 3 месяца назад
Отправлено 1 сентября 2009 г. в 7:37
JasperReports не умеет суммировать java.sql.Timestamp значения. Обратите внимание, что концептуально java.sql.Timestamp — это момент времени, а не длина временного интервала, поэтому суммирование таких значений не обязательно имеет смысл.
Но, как вы сказали, суммирование значений java.lang.Long возможно, и отображение длинного значения в качестве метки времени не должно быть трудным. См. Образец ниже.
С уважением,
Люциан
| Код: |
|
Присоединился: 17 июля 2006 г. — 1:10
Последний визит: 1 месяц 1 неделю назад
Отправлено 1 сентября 2009 г. в 13:49
Это сработало.Спасибо, Люциан ..
Зарегистрирован: 2 апреля 2009 г. — 17:03
Последний визит: 12 лет 3 месяца назад
Анализ транскрипционной логики, управляющей дифференциальной пространственной экспрессией в генах-мишенях Hh
Abstract
Эта работа предоставляет теоретические инструменты для анализа транскрипционных эффектов определенных биохимических механизмов (т.е. affinity and cooperativity), которые были предложены в предыдущей литературе для объяснения правильной пространственной экспрессии генов-мишеней Hedgehog, участвующих в развитии Drosophila . В частности, мы сосредоточились на экспрессии decapentaplegic , wingless , stripe и patch . Считается, что транскрипция этих генов контролируется модулями энхансеров, способными интерпретировать противоположные градиенты активаторной и репрессорной форм фактора транскрипции Cubitus interruptus (Ci).Это исследование основано на термодинамическом подходе, который обеспечивает уровни экспрессии этих генов. Эти скорости экспрессии контролируются факторами транскрипции, которые конкурируют и взаимодействуют за общие сайты связывания. Мы сделали математические представления различных показателей экспрессии, которые зависят от множества факторов и переменных. Выражения, полученные с помощью модели, были уточнены для получения более простых эквивалентных формул, которые позволяют проводить их математический анализ. Благодаря этому мы можем оценить корреляцию между различными взаимодействиями, участвующими в транскрипции, и биологическими особенностями, наблюдаемыми на тканевом уровне.Эти математические модели могут быть применены к другим морфогенам, чтобы помочь понять сложную транскрипционную логику противоположных градиентов активатора и репрессора.
Образец цитирования: Cambón M, Sánchez Ó (2019) Анализ транскрипционной логики, управляющей дифференциальной пространственной экспрессией в генах-мишенях Hh. PLoS ONE 14 (1): e0209349. https://doi.org/10.1371/journal.pone.0209349
Редактор: Сюзанна Резерфорд, Онкологический исследовательский центр Фреда Хатчинсона, США
Поступила: 05.06.2018; Одобрена: 4 декабря 2018 г .; Опубликовано: 7 января 2019 г.
Авторские права: © 2019 Cambón, Sánchez.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: Эта работа частично поддержана: Ministryio de Economía y Competitividad (Правительство Испании) по исследовательским грантам MTM2014-53406-R и FPI2015 / 074837 http: // www.mineco.gob.es/portal/site/mineco/ и Consejería de Economía, Innovación, Ciencia y Empleo, Проект Хунта де Андалусия (правительство Андалусии) FQM 954, https://www.juntadeandalucia.es/organismos/empleoempresayrcomerml . Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Hedgehog (Hh) — это морфоген, сигнальный белок, который вызывает несколько клеточных ответов.Он участвует в развитии различных биологических систем, например, у мухи Drosophila melanogaster . В имагинальном диске крыла Drosophila секреция Hh из клеток заднего компартмента индуцирует экспрессию нескольких генов-мишеней внутри клеток переднего компартмента. Среди них decapentaplegic ( dpp ) и исправленный ( ptc ). Оба вызывают синтез соответствующих им белков, Dpp и Ptc, которые важны для развития центрального домена крыла [1, 2].В эмбриональной эктодерме Hh также регулирует гены wingless ( wg ) и stripe ( sr ).
Однако известно, что один и тот же сигнал Hh вызывает разную пространственную экспрессию этих генов. То есть экспрессия ptc ограничена только дисковыми зонами, близкими к границе Anterior / Posterior (A / P) с высокими концентрациями Hh, в то время как dpp экспрессируется в более широком диапазоне диска при низких концентрациях Hh. .Возникает вопрос: почему один и тот же сигнал вызывает разные пространственные выражения для разных генов? Ответ на этот вопрос все еще обсуждается. В настоящее время считается, что оба гена в основном отвечают одним и тем же принципам, которые мы перечисляем ниже.
Hh транскрипционно контролирует как Dpp, так и Ptc через факторы транскрипции (TFs) Cubitus interruptus (Ci). Он определяет активность ферментов РНК-полимеразы (РНКП), которые контролируют генетическую транскрипцию посредством синтеза рибонуклеиновой кислоты (мРНК).Этот процесс требует связывания RNAP с некоторыми конкретными сайтами в цепи ДНК, называемыми промоторами. Однако скорость транскрипции генов-мишеней не только зависит от общей концентрации РНКП в системе, но также контролируется белком Ci. Ci присутствует в двух противоположных формах: активатор и репрессор. Активаторы CiA пытаются повысить скорость транскрипции, в то время как репрессоры CiR пытаются ее снизить. Сигнал Hh влияет на баланс между обеими формами Ci, т.е. в отсутствие Hh Ci появляется в подавленной форме, но когда Hh поглощается клеткой, Cubitus меняет свою роль, представляя свою форму активатора.Таким образом, градиент Hh в переднем отделе создает противоположные градиенты активатора (CiA) и репрессора (CiR). Более того, обе формы Ci должны связываться со специфическими сайтами ДНК, называемыми энхансерами или цис-регуляторными сайтами, которые отличаются от сайтов связывания RNAP. Один промотор может регулироваться одним или несколькими относительно короткими модулями энхансера, которые активируются / репрессируются связыванием нескольких TF. Таким образом, паттерн экспрессии гена отражает совокупную активность всех модулей энхансера, которые способны активировать / репрессировать его транскрипцию [3, 4].В этой работе мы планируем проанализировать контроль, выполняемый конкретным типом модулей, названных целевыми энхансерами Hh / Ci, которые объединяют конкурирующие входные данные с противоположными функциями транскрипции [5]. Только ограниченное количество прямых энхансеров-мишеней Hh / Ci было идентифицировано у Drosophila за последнюю четверть века в генах-мишенях Hh как dpp , ptc , wg или sr , подробности и другие примеры см. Таблица 1 в [5], [6] и ссылки в них.Среди них мы видим, что dpp в имагинальном диске Drosophila одновременно активируется и репрессируется CiA и CiR, которые постоянно конкурируют за связывание модуля из 3 энхансеров ( dppD ) [7, 8] .
Мы должны отметить, что контроль этих модулей Hh / Ci, вероятно, не оправдывает абсолютную генетическую экспрессию целевого гена, хотя они были описаны как существенные для их соответствующих генетических пространственных паттернов [5]. Кроме того, методология и результаты, представленные в этой работе, позволяют нам понять вклад одного модуля Hh / Ci даже в отсутствие знаний о контроле, выполняемом другими факторами транскрипции или другими модулями энхансеров.
В некоторых недавних работах [5, 9–11] постулируется, что причина правильной пространственной экспрессии этих генов может быть найдена в определенных биохимических факторах, участвующих в процессе транскрипции. Во-первых, связывание как РНКП, так и Ci в промоторе и энхансерах осуществляется посредством химических реакций. Для этого требуется некоторая свободная энергия, которая обычно характеризуется величиной, называемой сродством связывания. Это сродство зависит от нескольких характеристик промоторов и энхансеров каждого транскрибируемого гена.Фактически, в [5] было замечено, что энхансеры с более низким относительным сродством, по-видимому, необходимы для получения нормальной экспрессии dpp в областях со слабым сигналом. Во-вторых, возможно, что факторы транскрипции, которые уже связаны в некоторых энхансерах, могут изменять сродство других связывающих элементов. В этом случае связанные TF могут изменять свободную энергию последующей реакции связывания либо TF, либо RNAP. Этот процесс обычно называют кооперативностью, однако он может быть положительным или отрицательным.Если это способствует связыванию, это называется (нормальной) кооперативностью, а если это препятствует, — антикооперативностью. В [12, 13] было высказано предположение, что TF активатор / репрессор изменяют скорость транскрипции, стимулируя или блокируя соответственно рекрутирование РНК-полимеразы. Это означает, что кооперативность или антикооперативность с РНКП изменяет аффинность связывания промотора. Комбинация всех этих биохимических факторов (конкуренция, кооперативность и аффинность связывания) приводит к очень сложному балансу между концентрацией активаторов и репрессоров, что затрудняет различение их взаимодействующих эффектов на тканевом уровне.
В [5, 10] пространственная экспрессия некоторых генов-мишеней Hh была связана с соответствующей аффинностью связывания между белками Cubitus и энхансерами модуля Hh / Ci. Относительное сродство in vitro сайтов Ci в энхансерах ptc и dpp было измерено с помощью анализов сдвига электрофоретической подвижности, см. Рис. S2 в [10]. На рис. 4A в [11] аффинность связывания Ci четырех сайтов Ci в энхансере эмбриональной эктодермы wg была измерена с использованием той же методологии.В Таблице 1 в [5] сайты связывания Ci ранжируются для 9-мер в порядке предсказанной аффинности связывания с Ci для генов в геноме Drosophila , для которых Hh / Ci-регулируемые энхансеры были функционально охарактеризованы. Было замечено, что ptc активируется Hh / Ci в имагинальных дисках личинок через модуль с высокоаффинными Ci сайтами, в отличие от энхансеров dpp , wg и sr с относительно низким сродством. расположены в соответствующих модулях Hh / Ci.
Эксперименты, проведенные в [10], подтверждают, что при умеренном сигнале Hh сайты с низким сродством дикого типа в dppD вызывают активацию, тогда как при замене сайтов с высоким сродством происходит репрессия. Аналогичные результаты были получены для эмбриональных энхансеров wg и sr в [5, 11]. Отметим, что эта активация / репрессия относится к уровням транскрипции, наблюдаемым в отсутствие какого-либо энхансера в модуле Hh / Ci (рис. 2 в [10], дополнительный рис. 5 в [11], рис 2E в [5] и [14]).Ясно существование др. Механизмов для интерпретации градиента Hh в остальной части локуса. В частном случае dpp уровни экспрессии в отсутствие этих трех сайтов связывания в dppD показывают глубокую репрессию вблизи границы A / P. Насколько известно автору, это подавление до конца не изучено (см. Результаты и обсуждение (h) в [5]). Тем не менее, вклад (активация / репрессия) единственного модуля энхансера dppD можно оценить с помощью анализа относительных уровней экспрессии.
Для того, чтобы различать механизмы, которые могут вызвать дифференциальное пространственное выражение, в [10, 11] эксперименты были сопоставлены с приспособлениями к термодинамической модели, основанной на идеях Ши, Аккерса и соавторов [15, 16]. Более того, подгоняя модель кооперативности репрессоров в [10], они обнаружили, что CiR играет существенную роль в ответе на умеренный сигнал Hh. В [11] авторы также предположили, что кооперативность между репрессорами может играть важную роль в изменении генетической экспрессии вдоль имагинального диска, с помощью математической модели конкуренции за занятость между репрессорами и активаторами.
Большое количество биохимических переменных, присутствующих в системе, требует математических моделей [17, 18], которые могут пролить некоторый свет на происхождение дифференциальной пространственной экспрессии в генах-мишенях Hh, среди прочего. Термодинамическая модель, предложенная Shea, Ackers и соавторами [15, 16], также известная как BEWARE [19] (Binding Equilibrium Weighted Average Rate Expression), является методом, часто используемым в математическом моделировании процессов генетической транскрипции. См. [20] или [21] для общего обсуждения / сравнения с другими подходами к моделированию, например с булевыми моделями.Однако эта модель порождает длинные и сложные математические выражения, даже если задействовано всего несколько факторов транскрипции. Для анализа независимых и специфичных сайтов связывания и анализа двух неконкурентных факторов транскрипции ранее были предложены только простые математические выражения [22, 23]. Трудно расшифровать биологические эффекты в модели, даже если они поддерживаются численными инструментами [24], потому что выражения по своей сути включают большое количество констант и переменных.
В этой работе мы пытаемся лучше понять транскрипционную логику генов-мишеней, контролируемых модулем Hh / Ci энхансеров, с теоретической точки зрения, используя термодинамическую модель. Наш анализ предполагает, что транскрипционная логика в присутствии противоположных градиентов активатор / репрессор может демонстрировать разные версии в зависимости от кооперативности между факторами транскрипции. Фактически, разработанная нами теоретическая методология способна продемонстрировать, как сочетание схожих биохимических факторов, применяемых в различных рамках, вызывает совершенно разные транскрипционные эффекты.Мы протестировали эту общую схему с экспериментальными результатами для нескольких генов-мишеней Hh, таких как dpp , ptc , wg и sr . Среди всех выведенных структур мы делаем вывод, что правильная пространственная экспрессия этих генов обусловлена различиями в аффинности между их соответствующими сайтами связывания в сочетании с кооперативностью между репрессорами (для получения более подробной информации обратитесь к разделу «Результаты»). Наш анализ подтверждает результаты, полученные в [4, 5, 10, 11].
Однако это не исключает, что управление, выполняемое модулем Hh / Ci, может модулироваться другими модулями энхансера или сигналами в зависимости от биологического контекста. Недавно в [4] было описано, что активность модуля Hh / Ci может быть специфичной для стадии и ткани, что может быть связано с синергетической работой различных модулей.
В случае градиента одного активатора наш анализ представляет хорошо принятую транскрипционную логику, которая исходит из пороговой модели активатора .Эта модель объясняет роль определенных биохимических факторов, участвующих в интерпретации сигналов. Например, различное сродство активаторов к элементам ДНК [25] и кооперативность между активаторами [10, 26]. Сайты связывания с высоким сродством и кооперативность между активаторами благоприятствуют связыванию активаторов с энхансерами, обеспечивая экспрессию генов при низких концентрациях активатора, поэтому здесь мы наблюдаем более широкий ответ в градиенте активатора. Напротив, сайты с низким сродством и отсутствие кооперативности между активаторами ограничивают экспрессию гена областями с высокой концентрацией активатора.Хотя это обоснование хорошо принимается в сценарии единственного градиента, оно не было успешно применено в комбинаторных взаимодействиях, как, например, в противоположных градиентах активатор-репрессор, таких как те, которые мы описали в передаче сигналов Hh [10, 25]. В этом случае баланс между обоими градиентами вызывает существование диапазонов чистых активированных / репрессированных клеток, т.е. клеток вдоль ткани, которые экспрессируют более высокие / более низкие уровни, чем базальный уровень [10], и, следовательно, нет глобальной активации / репрессии.Клеточные диапазоны экспрессии (CER) будут определяться не только интенсивностью сигнала, но также чистыми активированными / репрессированными клеточными диапазонами.
Наш анализ предполагает, что биохимические различия между генами могут влиять как на модуляцию сигнала, так и на изменения в чистом активированном диапазоне клеток. То есть вариацию ССВ можно объяснить путем анализа комбинации обоих аспектов. Очень примечательным моментом в этой работе является то, что использование одних и тех же биохимических характеристик с разной кооперативностью между активаторами и репрессорами даст совершенно разные скорости экспрессии.Различные типы кооперативных взаимодействий между факторами транскрипции будут приводить к вариациям в логике транскрипции. Еще один очень интересный аспект нашего анализа — это то, как количество энхансеров может изменять экспрессию гена. В [10] была сконструирована линия трансгенной мухи, несущая репортер GFP с одним высокоаффинным сайтом Ci, чтобы определить, играет ли кооперативность между TF важную роль в процессе передачи сигналов. Даже при отсутствии кооперативности такое изменение количества энхансеров могло изменять экспрессию гена, по крайней мере, теоретически.Этот аспект будет играть ключевую роль в нашем аргументе для понимания генов-мишеней Hh. Хотя они, по-видимому, играют центральную роль, сродство и кооперативность между TF не являются единственными биохимическими факторами, участвующими в интерпретации общей передачи сигналов морфогенов. В развитии нервной трубки эмбриона цыпленка / мыши, другого парадигматического примера формирования морфогенетического паттерна, клетки дифференцируются в ответ на морфогенетические сигналы Sonic Hedgehog (Shh) [27-30]. В этом случае сигнал Shh уравновешивает концентрацию различных версий активаторов и репрессоров семейства Gli.Эти Gli TF распознают последовательности-мишени, которые очень похожи, однако было высказано предположение, что некоторые TF обладают большей эффективностью в качестве активаторов или репрессоров [26], чем другие. Таким образом, различная эффективность активаторов и репрессоров может быть еще одним фактором, который следует принимать во внимание при использовании дифференциальной кооперативности TFs-RNAP.
Результаты
В этой работе мы изучим, какие из предыдущих факторов могут изменять диапазоны клеточной экспрессии, анализируя их комбинированное влияние как на модуляцию сигнала, так и на изменение диапазонов чистых активированных / репрессированных клеток.Мы начнем с упражнения по моделированию, которое предоставит математическое представление (операторы) уровней выражения для последующего анализа. Поскольку нас интересует транскрипционный эффект модуля Hh / Ci, мы применяем метод BEWARE, чтобы получить уровни экспрессии гена, контролируемые двумя противоположными общими факторами транскрипции: активатором CiA и репрессором CiR. Полученные «осторожные операторы» включают для каждого поколения:
- факторов транскрипции, конкурирующих за связывание одного из n общих цис-регуляторных сайтов,
- отдельных аффинностей связывания ТФ в зависимости от их активаторной или репрессорной формы,
- взаимодействий кооперативности между TF и кооперативности / антикооперативности между TF и RNAP.Мы рассматриваем различные процессы кооперативности TF-TF, которые приводят к различным выражениям оператора BEWARE:
- отказ от сотрудничества,
- общая кооперативность, когда связанный TF изменяет аффинность связывания любого другого фактора транскрипции,
- частичная кооперативность, которая имеет место только между TF, которые имеют одну и ту же форму, активаторы взаимодействуют только с активаторами, а репрессоры взаимодействуют только с репрессорами.
Панель A на фиг. 1 показывает все эти взаимодействия с n = 3 сайтами связывания.Мы упростили математические выражения операторов BEWARE, поскольку в их исходной форме невозможно было бы провести последующий математический анализ.
Рис. 1. Сеточно-активированный клеточный (NAC) диапазон, описанный термодинамической моделью.
A) Схема эксперимента по определению диапазона NAC. Стрелки представляют все возможные взаимодействия, зафиксированные термодинамической моделью, определяющей скорости транскрипции: двуглавые прямые стрелки показывают аффинности сайта связывания белок-ДНК, тогда как одинарные черные и красные стрелки обозначают кооперативность TFs-RNAP и TFs-TF соответственно.Чистый активированный клеточный диапазон dpp D3xCi ptc , репортерного гена с версией энхансера dpp с тремя высокоаффинными сайтами связывания, получают путем сравнения его теоретической транскрипционной активности с активностью dpp. D3xCi KO , ген, содержащий другую версию энхансера dpp , содержащую три сайта с нулевым сродством. Оба случая представлены на верхней и нижней схемах соответственно.Сайты связывания TF представлены закругленными прямоугольниками, закрашенными зеленым (высокое сродство) или черным (нулевое сродство). B) Теоретические скорости транскрипции, предсказанные для обоих генов в клетках переднего компартмента. Этот отсек занимает 60% имагинального диска Drosophila , а задний отсек — остальное (от 60% до 100%). Уровни экспрессии, задаваемые операторами BEWARE, составляют от 0 нМ / мин до 1 нМ / мин , что является базальным уровнем, равным 0.5 нМ / мин . Эти эталонные уровни экспрессии были выбраны для правильной оценки модуляции сигнала. Поскольку dpp D3xCi KO был смоделирован независимо от внешних факторов, он выражается на базальном уровне где угодно. Клетки, экспрессирующие dpp D3xCi ptc больше, чем базальный уровень, находятся в диапазоне NAC. Экспрессия обоих генов в имагинальном диске крыла также указывается с помощью цветных полос.Синий кружок внутри полосы указывает положение клетки, экспрессирующей dpp D3xCi ptc на базальном уровне. Цветовая шкала, используемая в этих столбцах, показана в C) , черный означает отсутствие выражения (0 нМ / мин ), а полный цвет означает высокое выражение (1 нМ / мин ). На вставке в B) изображены градиенты активатор / репрессоры (CiA / CiR), генерируемые передачей сигналов Hh: концентрации активатора выше вблизи передней / задней границы.Более подробное описание можно найти в уравнении (16).
https://doi.org/10.1371/journal.pone.0209349.g001
Анализ этих математических выражений позволяет оценить, какие биохимические характеристики могут изменять пространственную экспрессию двух генов, контролируемых одними и теми же ТФ, и каким образом . Экспериментальные доказательства, которые мотивируют наш анализ, в основном связаны с аффинностью и числом n сайтов связывания. С помощью анализа сдвига электрофоретической подвижности Паркер и соавторы обнаружили в [10], что сайты связывания Ci в энхансере ptc имеют значительно более высокое сродство, чем сайты dpp .Те же авторы сконструировали трансгенные линии мух, которые позволяют им сравнивать транскрипционную активность репортерных генов, содержащих разные варианты этих сайтов, модифицирующих их аффинность. Подобные результаты были описаны у эмбриона для wg в [11].
Поскольку есть два противоположных сигнала, мы должны определить, когда клетка активирована или подавлена. Эти представления были заимствованы из [5, 10], где при использовании репортерных генов активность различных версий энхансера dpp , содержащих три низкоаффинных сайта ( dpp D-Ci WT ), три высоких — Сравнивали сайты аффинности ( dpp D-3xCi ptc ) или три сайта нулевой аффинности ( dpp D-3xCi KO ).Репортерный ген с сайтами с нулевым сродством обеспечивает базальную экспрессию, поскольку он отражает влияние всех других факторов, отличных от Ci, на модуль dppD. Мы отсылаем к результатам и обсуждению (h) в [5] для более подробного обсуждения. В частности, в [5, 10] были измерены эффекты передачи сигналов Ci с помощью энхансеров с низким или высоким сродством, сравнивая активность гена с базальной активностью в любой клетке. Клетки, экспрессирующие ген с более высокими показателями экспрессии, чем базальный уровень, называются чистыми активированными клетками.Набор всех чистых активированных клеток составляет диапазон чистых активированных клеток (NAC). На рис. 1 показано, как этот диапазон определяется с помощью термодинамической модели таким же образом, как это было сделано из измерений в [10].
Мы хотим выяснить, какие концентрации активаторов и репрессоров, [CiA] и [CiR], будут обеспечивать большую или меньшую экспрессию генов, чем базовая. Итак, мы определяем пороговое значение, разделяющее концентрации обоих TF, которые будут производить чистые активированные клетки или чистые репрессированные клетки, используя операторы BEWARE.Затем мы можем использовать порог между чистыми концентрациями активации / репрессии, чтобы определить предел между диапазонами чистых активированных или репрессированных клеток. После того, как мы определили диапазоны активированных клеток, мы можем предсказать, как биохимические различия повлияют на них, а также на интенсивность сигнала. Для проведения этого анализа мы должны предположить, что противоположные градиенты активатора и репрессора являются монотонными и не меняются со временем (см. Панель B на Рис. 1 для графического примера). Отметим, что применяемый нами подход не зависит от конкретных значений концентраций TF.Математический анализ может определить, как биохимические различия вызывают вариации скорости транскрипции, только предполагая, что они реагируют на одни и те же (неизвестные) противоположные концентрации активатора / репрессора. Это контрастирует с большой вариабельностью в определении градиентов TF, демонстрируемой теоретическими моделями, подобранными в [10]. Эта методология подробно описана в разделе «Методы». Отметим, что другой теоретический подход был предложен в предыдущей работе [11].
С помощью проведенного анализа мы определили, какие биохимические факторы могут участвовать в дифференциальной экспрессии, наблюдаемой в генах-мишенях Hh.Для этого мы тестируем и сравниваем наши теоретические результаты с уже существующими экспериментальными данными, в частности, для dpp , wg , sr и ptc . Во всех случаях мы сначала описываем эксперименты и наблюдаемые результаты. На втором этапе мы сравниваем эти измерения с различным качественным поведением, предсказанным нашим моделированием, и, наконец, мы выводим биохимическую основу, которая объясняет экспериментальные данные. Сначала мы применим эту методологию к dpp и модулю расширения dppD.В этом случае существует несколько независимых экспериментальных подходов, предсказывающих одну и ту же биохимическую основу, которые можно интерпретировать как подтверждение нашего теоретического анализа.
Эксперимент 1: Транскрипционные эффекты восстановительных сайтов связывания
В [10] трансгенная линия мух, несущая репортер GFP с одним высокоаффинным сайтом Ci ( dpp D-1xCi ptc , где верхний индекс «ptc» обозначает высокоаффинную версию dpp энхансеры в соответствии с исходной номенклатурой).В этом случае диапазон чистых активированных клеток был шире, чем диапазон чистых активированных клеток для ( dpp D-3xCi ptc ), и, как следствие, более широкая, но ослабленная экспрессия наблюдалась для случая одного энхансера. чем в случае с 3 сайтами с высоким сродством. Это сравнение можно воспроизвести с помощью операторов BEWARE. Теоретически можно доказать, что кооперативность между ТФ определяет эффект уменьшения сайтов связывания:
- При наличии полной кооперативности или некооперативности между TF диапазон NAC по существу останется неизменным, хотя может наблюдаться снижение интенсивности сигнала, то есть меньшая репрессия / активация в репрессированных / активированных клетках.
- Если активаторы CiA взаимодействуют только между собой, то будет наблюдаться сокращение диапазона NAC, поэтому некоторые чистые активированные клетки для dpp D-3xCi ptc будут изменены, чтобы быть чистыми репрессированными для гена dpp D- 1xCi ptc .
- Наконец, в случае кооперативности только между репрессорами диапазон NAC будет увеличен в соответствии с измерениями для dpp D-3xCi ptc и dpp D-1xCi ptc , полученных в [ 10].
Эти результаты (суммированные в таблице 1, строка 3) ) уравновешивают двоякое следствие уменьшения количества энхансеров с 3 до 1. С одной стороны, уменьшение подразумевает исчезновение любого возможного вида кооперативности между TF. . В случае полной кооперативности между ТФ эти отношения симметричны для активаторов и репрессоров, поэтому их исчезновение снижает передачу сигналов, то есть меньше транскрипции в активированных клетках и больше транскрипции в репрессированных клетках, но это не изменяет баланс между чистыми активированными или репрессированными. сотовые диапазоны.В случае асимметричной кооперативности, то есть частичной кооперативности только между активаторами или репрессорами, кооперативная разновидность теряет это преимущество. Это может спровоцировать глобальное снижение активации в случае кооперативности активаторов и репрессии в случае кооперативности репрессоров. Это вызывает изменение диапазона NAC: когда кооперативность между активаторами удаляется, диапазон NAC уменьшается и увеличивается, когда кооперативность репрессора отменяется. Интерпретация этих утверждений, напротив, позволяет нам утверждать, что одна из ролей асимметричной кооперативности между активаторами / репрессорами заключается в увеличении / уменьшении диапазона NAC по сравнению с диапазоном NAC в некооперативном случае.Сводку этих результатов можно найти в таблице 1, строка 2) . С другой стороны, независимо от кооперативности, уменьшение количества энхансеров также означает, что передача сигналов должна быть ослаблена. Оба соображения объясняют теоретические транскрипционные эффекты уменьшения сайтов связывания.
На рис. 2 представлены конкретные примеры различного поведения в диапазоне NAC в рамках одного и того же эксперимента в присутствии ранее упомянутых взаимодействий. Согласно нашему моделированию, единственная транскрипционная логика, совместимая с экспериментами, — это та, которая возникает в присутствии частичной кооперативности между репрессорами.Тем не менее мы можем найти больше совпадений в этом направлении, используя другие экспериментальные данные.
Рис. 2. Транскрипционные ответы на эксперимент 1.
Первый столбец: схема эксперимента 1: сравнение диапазонов экспрессии репортерных генов с 3 сайтами с высоким сродством ( dpp D-3xCi ptc ) и единственный сайт Ci с высоким сродством ( dpp D-1xCi ptc ). A) соответствует случаю не / полной кооперативности, где, если кооперативность сохраняется, все TF взаимодействуют между ними, C) к случаю кооперативности активаторов, кооперируются только активаторы, и, наконец, E) к случаю кооперативности репрессоров где сотрудничают только репрессоры.Во втором столбце показаны различные транскрипционные ответы, которые могут быть теоретически описаны в зависимости от рассматриваемого случая кооперативности. В схемах и графиках используются те же ключи, что и на рис. 1.
https://doi.org/10.1371/journal.pone.0209349.g002
Эксперимент 2: эффекты дифференциального сродства
Второй эксперимент, на котором мы сосредоточимся, — это сравнение чистых скоростей транскрипции репортерных генов, содержащих либо три версии энхансера с высоким сродством dpp ( dpp D-3xCi ptc ), либо три варианта с низким сродством dpp сайтов ( dpp D-Ci WT ).Было замечено, что более высокая аффинность Ci обеспечивает снижение чистой активированной клеточной области, которая является релевантной промежуточной областью, где чистые активированные клетки для ( dpp D-Ci WT ) репрессируются для ( dpp D). -3xCi ptc ). Также было замечено, что повышенное сродство обеспечивает более сильную активацию в области, близкой к границе A / P, а также более сильную репрессию в областях, далеких от той же границы (см. Рис. 2D в [10]).
Если мы примем, что кооперативность между ТФ работает одинаково при связывании с обоими, dpp и ptc , версиями сайтов связывания, наш анализ предполагает, что эффект снижения аффинности может снова зависеть от тип взаимодействия между ТФ:
- При наличии полной кооперативности или некооперативности между TF диапазон NAC по существу останется неизменным, хотя может наблюдаться снижение интенсивности сигнала, то есть меньшая репрессия / активация в чистых репрессированных / активированных клетках.
- Если активаторы CiA взаимодействуют только между собой, диапазон NAC будет уменьшен, поскольку можно доказать, что этот диапазон монотонно возрастает с увеличением аффинности, то есть чем больше сродство, тем шире диапазон NAC.
- В случае кооперативности только между репрессорами диапазон NAC будет увеличиваться, потому что этот диапазон монотонно уменьшается с увеличением аффинности, то есть чем больше аффинность, тем уже диапазон NAC.
И снова кооперативная модель репрессоров является единственной, согласующейся с результатами, наблюдаемыми для dpp D-3xCi ptc и dpp D-3xCi WT в [10].Это теоретическое поведение можно увидеть на рис. 3. Во всех предыдущих анализах, суммированных в таблице 1 строка 1) , считалось, что изменение аффинности энхансеров пропорционально влияет как на активаторы, так и на репрессоры. См. Подробности в разделе «Методы».
Рис. 3. Транскрипционные ответы на эксперимент 2.
Рисунки в первом столбце: схема эксперимента 2: сравнение диапазонов экспрессии репортерных генов с 3 сайтами с высоким сродством dpp D-3xCi ptc или 3 сайта с низким сродством dpp D-3xCi WT .Аффинности сайтов связывания TFs-ДНК указаны более толстыми или более тонкими двуглавыми прямыми стрелками. A) соответствуют случаю отсутствия кооперативности или полной кооперативности, когда, если кооперативность сохраняется, все TF взаимодействуют между ними, C) к случаю кооперативности активаторов, кооперируются только активаторы, и, наконец, E) к случаю кооперативности репрессоров где сотрудничают только репрессоры. Цифры во втором столбце показывают транскрипционные ответы, которые могут быть теоретически описаны в зависимости от рассматриваемого случая кооперативности.В схемах и графиках используются те же ключи, что и на рис. 1.
https://doi.org/10.1371/journal.pone.0209349.g003
Таким образом, в случае dpp оба результата совместимы только при наличии сотрудничества между репрессорами. Этот вывод совпадает с выводом, полученным в [10] с использованием числовой подгонки к экспериментальным данным, и с [5], где авторы обнаружили, что dpp нуждается в сайтах связывания с низким сродством для нормальной активации в областях с низкой передачей сигналов Hh.
Эксперимент 2 также был проведен на эмбрионе Drosophila для wg и sr с заменой модулей энхансеров Hh / Ci с низким относительным сродством на высокое сродство (см. Дополнительный рисунок 5 в [11], рисунок 2E в [ 5] и [14]). Было обнаружено, что увеличение сродства Ci в соответствующих энхансерах ограничивает экспрессию. Наше моделирование предполагает, что в обоих случаях должна быть задействована кооперация репрессоров. Более того, в случае wg увеличение сродства сайтов Ci вызывает только репрессию.Этот факт также представлен нашей моделью (см. Панель A на фиг. S3, где клетки, содержащие концентрации Ci между красной и зеленой кривыми, будут меняться с чисто активированных на чистые репрессированные, если сродство энхансеров увеличивается). Наконец, мы собираемся проанализировать совместимость наших результатов с диапазонами экспрессии ptc . Известно, что cubitus interruptus имеет более высокое сродство к энхансерам ptc , чем dpp ’s, см. Рис. S2 в [10]. Однако эксперименты также показали, что экспрессия в таких модулях, по-видимому, регулируется исключительно активаторной формой CiA.В частности, в [8] они трансплантировали Ci-связывающий домен гена ptc в контекст энхансера dpp . В своих результатах они наблюдали, что, в отличие от его нормальной роли, фрагмент ptc в своем новом окружении опосредует реакцию на CiR. Мотивированные этим экспериментом, мы предположили в нашей модели, что ptc не только управляется одиночным градиентом CiA, но и противоположными градиентами cubitus, где репрессорная форма является очень слабым репрессором транскрипции.С помощью этой дополнительной гипотезы мы пришли к выводу, что частичная кооперативность между CiR и высокой аффинностью связывания снова является каркасом, совместимым с тем фактом, что ptc обнаруживает более короткую CER в развитии имагинального диска крыла Drosophila .
Действительно, наш анализ позволяет нам интерпретировать роли дифференциальной аффинности и репрессорной кооперативности для генов-мишеней Hh в следующих терминах:
- Анализ эксперимента 1 показывает нам, что кооперативность репрессоров снижает диапазоны NAC обоих генов по отношению к случаю некооперативности.
- Кроме того, анализ эксперимента 2 предполагает, что это уменьшение области NAC менее эффективно с сайтами связывания с низким сродством dpp , чем с сайтами связывания с высоким сродством ptc . Даже, в зависимости от баланса между концентрациями активатора-репрессора, область NAC может исчезнуть, как это наблюдалось для wg .
Более того, предыдущая точка зрения будет означать, что диапазон NAC для dpp D-1xCi ptc должен соответствовать диапазону NAC для dpp D-3xCi WT , а последняя должна содержат диапазон NAC для dpp D-3xCi ptc .Фактически, эти соотношения полностью совместимы с результатами [10]. Более конкретно, на рисунках 2 и 4 в [10] можно заметить, что диапазоны NAC для dpp D-1xCi ptc , dpp D-Ci WT и dpp D- 3xCi ptc занимают от 43%, 49% и 54% ширины диска соответственно до границы A / P (что составляет примерно 60% ширины диска).
Логика транскрипции при наличии противоположных факторов транскрипции
Хотя результаты нашего анализа ранее применялись непосредственно к генам-мишеням Hh, анализ может быть выполнен для любых других генов, контролируемых противоположными факторами транскрипции, A и R, активаторами и репрессорами соответственно.С этого момента мы будем применять этот общий подход, и в частном случае генов-мишеней Hh CiA и CiR будут играть роль A и R.Как было указано во введении, предыдущие результаты несовместимы с транскрипционной логикой пороговая модель активатора. Итак, в этом разделе мы опишем варианты транскрипционной логики, которые могут быть найдены в зависимости от кооперативности между TF: не / полная кооперативность, частичная кооперативность активаторов и частичная кооперативность репрессоров.
Результаты нашего анализа суммированы в Таблице 1. Она показывает относительный размер диапазонов клеточной экспрессии (CER) двух генов, gene1 и gene2 , которые контролируются одинаковыми противоположными градиентами TF, хотя и демонстрируют некоторые различия в их значениях. биохимические характеристики. Рассмотренные биохимические различия между gene1 и gene2 перечислены в строках:
- ↓ Сродство ТФ: сродство к сайтам связывания гена 1 меньше, чем сродство тех же ТФ к сайтам связывания гена 2 .Следуя [10], это происходит, например, в случае dpp D-3xCi WT и dpp D-3xCi ptc . Рассматриваемое уменьшение сродства пропорционально для A и R, .
- ↓ ТФ-ТФ кооперируются: ТФ взаимодействуют менее интенсивно при связывании с энхансерами гена1 , чем при связывании с энхансерами гена2 , но оба гена имеют одинаковый тип кооперативности,
- ↓ нет. энхансеры: gene1 имеет меньше сайтов связывания, чем gene2 , как, например, гены dpp D-1xCi ptc и dpp D-3xCi ptc ,
- ↓ A-RNAP кооператив.: активатор A является более слабым активатором транскрипции для gene1 , чем для gene2 , проявляя более слабую кооперативность между активаторами и РНК-полимеразой,
- ↓ R-RNAP coop: репрессор R является более слабым репрессором транскрипции для gene1 , чем для gene2 , проявляя более слабую антикооперативность между репрессорами и РНК-полимеразой.
Результаты, представленные в таблице 1, также суммируют способ, которым происходят изменения CER.↑ CER / ↓ CER указывают на то, что gene1 имеет более широкий / более узкий CER, чем gene2 соответственно. Эти вариации CER зависят от модуляции сигнала, а также от вариаций диапазона NAC. В этом смысле теоретическая модель предполагает, что могут возникнуть следующие ситуации:
- ↓ Действие: Глобальное снижение скоростей экспрессии gene1 по сравнению с gene2 ,
- ↓ Rep: Глобальное приращение скоростей экспрессии gene1 по сравнению с gene2 ,
- ↓ Sig: декремент передачи сигналов, то есть для gene1 более низкие скорости экспрессии в чистых активированных клетках и более высокие скорости экспрессии в чистых репрессированных клетках.
- ↓ NAC: уменьшение диапазона NAC, когда диапазон NAC для gene1 меньше, чем диапазон NAC для gene2 ,
- ↑ NAC: прирост чистого активированного клеточного диапазона gene1 по сравнению с диапазоном NAC gene2 ,
- NAC: когда диапазоны NAC для gene1 и gene2 равны.
Последствия различий в биохимических характеристиках, перечисленных в таблице 1, ясно оправдывают существование нескольких версий транскрипционной логики в присутствии противоположных градиентов в зависимости от типа кооперативности между TF.Область чистых активированных клеток очень важна в случае не / полной кооперативности, потому что эффекты некоторых изменений биохимических характеристик только ослабляют передачу сигналов (↓ Sig) и, как следствие, они не изменяют диапазон NAC (NAC) (см. Панели BD). на S1 рис.). В случае частичной кооперативности только между репрессорами или активаторами можно вывести общее увеличение или уменьшение скоростей экспрессии, которые явно вызывают увеличение или уменьшение чистой активированной клеточной области (см. Панель D на S2 и S3 фиг.).Однако не всегда изменение диапазона NAC происходит из-за глобального увеличения или уменьшения скоростей экспрессии, как это видно на панели B на S2 и S3, фиг. Тем не менее, мы можем наблюдать, что последствия, проистекающие из некоторых других биохимических характеристик, как те, которые описаны в Таблице 1, строки 4), 5) , качественно одинаковы, независимо от кооперативности. Таблица 1 была введена для ясности, хотя она не охватывает все отношения и свойства, выведенные в ходе нашего анализа.S1 и S3 Фигуры иллюстрируют результаты, указанные в строках таблицы 1 1), 2), 3) . На этих графиках пороги и скорости транскрипции представлены пурпурным для gene1 и зеленым для gene2 , чтобы оценить относительные различия.
Те же операторы BEWARE можно использовать для определения скоростей экспрессии для одного градиента активатора, просто установив на ноль концентрации репрессоров. Таким же образом мы можем проводить анализ экспериментов того же типа, и результаты согласуются с транскрипционной логикой пороговой модели активатора.См. Раздел K в файле S1.
Обсуждение
В этой работе мы пытаемся углубить текущее понимание генетической экспрессии. Используя теоретический подход, мы можем выделить биохимические механизмы, участвующие в экспрессии генов, которые действуют через противоположные градиенты TF активатора и репрессора, а также степень их участия. Более конкретно, модель предполагает, что TFs контролируют рекрутирование РНК-полимеразы, конкурируя за общие сайты связывания посредством процессов связывания, в которых могут быть задействованы различные аффинности и кооперативности.Это относится к конкретным модулям энхансеров в некоторых генах-мишенях Hh. Метод BEWARE — это хорошо зарекомендовавший себя инструмент моделирования, который позволяет нам представить тонкий баланс между противоположными сигналами с помощью математических выражений и увидеть, как на эти пропорции влияют другие участвующие биохимические характеристики. Сокращая ранее длинные формулы BEWARE в компактные математические выражения, мы можем сделать вывод о существовании нескольких различных форм логики транскрипции, то есть нескольких сценариев, в которых одни и те же биохимические характеристики генов приводят к совершенно различным последствиям на тканевом уровне.Подробное описание различных сценариев и взаимосвязи между ними позволяет нам противопоставить эту теоретическую основу свидетельствам, предоставленным конкретными экспериментами, модифицирующими процесс связывания TF. Это было достигнуто в случае генов-мишеней Hedgehog dpp , ptc , sr и wg , где мы получаем выводы, аналогичные тем, которые были получены в предыдущей работе с использованием других методов. Таким образом, мы можем получить информацию об относительном контроле транскрипции, осуществляемом обоими ТФ независимо от других, известных или неизвестных контролирующих веществ, которые могут действовать одновременно.
Эта работа расширяет применимость метода BEWARE, поскольку соответствующая качественная информация может быть извлечена из компактных моделей. Тот факт, что эти модели могут быть применены аналогичным образом к другим биологическим системам, означает, что есть много интересных выводов, выходящих за рамки данной статьи.
Методы
Удержание оператора BEWARE для двух противоположных градиентов TF
В качестве первого шага мы применяем идеи статистического термодинамического метода к гену g , контролируемому двумя противоположными факторами транскрипции { A , R }, активатором и репрессором соответственно.Наша цель здесь состоит в том, чтобы вывести выражения для изменения концентрации белка G с течением времени в терминах концентраций A и R, [ A ] и [ R ], т.е. (1) Здесь «BEWARE ()» представляет собой математическую функцию, определяющую зависимость относительно роли активации / репрессии TF. Это не зависит от других возможных факторов, имеющих отношение к эволюции белка, например, деградации или пространственной дисперсии. В модели реакции связывания TF и RNAP в энхансерах и промоторе, соответственно, намного быстрее, чем синтез белка G, поэтому он будет рассматриваться в термодинамическом равновесии, определяемом Законом действия масс.Если B представляет собой пустую регуляторную область, набор незанятых энхансеров-промотор, комплексы BA , BR и BRNAP имеют концентрацию в равновесии, определяемую где,, K RP , [ A ], [ R ] и [ RNAP ] — константы диссоциации и концентрации активаторов, репрессоров и РНК-полимеразы. Таким образом, коэффициенты и безразмерны. Верхний индекс (1) обозначает константу диссоциации реакции, которая происходит в отсутствие другого TF, ранее связанного с другим энхансером (обратите внимание, что, поскольку наборы имеют только один промотор, верхний индекс не нужен для константы диссоциации RNAP) .Заметим, что чем выше сродство между белком / комплексом и сайтом связывания, тем ниже константа диссоциации в соответствующей реакции связывания. Последовательное связывание более одного фактора транскрипции считается последовательным и конкурентным процессом, так что реакции задаются равновесными концентрациями здесь верхний индекс (2) обозначает константу диссоциации для реакции TF, который уже связывает оператора с одним TF в другом сайте. Конкуренция моделируется таким образом, что константа диссоциации конфигурации свободных сайтов не зависит от их положения, но может зависеть от других TF, уже связанных с тем же набором энхансеров посредством кооперативности или антикооперативности.
Мы обозначаем как некооперативные ТФ все те белки, сродство к энхансерам которых не модифицируется какими-либо ранее связанными ТФ, то есть они подтверждают и Это предположение подразумевает последовательную независимость равновесных концентраций, поскольку [ BRA ] = [ BAR ] . Правдоподобно предположить то же соотношение для более поздних связываний, то есть для j ≥ 2. Вследствие этой последовательной независимости мы обозначаем константы диссоциации как K A и K R без надстрочного индекса.Таким образом, если все рассматриваемые TF не кооперативны, мы легко делаем вывод, что концентрация в равновесии конфигурации с j A активаторами и j R связанными репрессорами, равна (2) независимо от последовательного порядка связывания и конкретных позиций, занимаемых TF. Хотя цис-регуляторные элементы дикого типа Drosophila включают в себя всего 3 сайта связывания, мы собираемся сравнить с экспериментами, в которых эти сайты связывания были уменьшены до 1.Таким образом, мы будем рассматривать в нашей модели n ≥ 1 количество сайтов связывания ТФ. Во всех случаях у нас есть ограничение на возможное количество связанных факторов транскрипции. Итак, j A + j R ≤ n должен быть проверен, и, как следствие, j 0 = n — j A — j R ≥ 0 обозначает количество свободных мест в конфигурации.
С другой стороны, кооперативность возникает, когда существование других ранее связанных белков влияет на сродство нового связывающего белка типа i , k = A , R , то есть: где c — положительная константа, больше 1, если белки взаимодействуют, и меньше 1, если происходит антикооперативность. Поскольку единственная разница между кооперативностью и антикооперативностью — это пороговое значение для c , в последующем моделировании мы будем ссылаться на константу c и не будем различать оба случая.Если кооперативность имеет место, необходимо знать, на какие TF влияют другие TF, поскольку равновесная концентрация будет зависеть от этих соотношений. В предыдущей литературе недавно было предложено, чтобы полная и частичная кооперативность играла важную роль в генах-мишенях Hh / Shh посредством Ci / Gli TFs [10, 11, 26]. Частичная кооперативность активаторов будет происходить, когда наличие связанного активатора одинаково изменяет аффинность связывания любого заднего активатора, то есть j ≥ 2.То же самое и с репрессорами. Полная кооперативность будет иметь место, когда присутствие связанного TF изменяет аффинность любого заднего связывания таким же образом, т.е. и одновременно для j ≥ 2 (см., Например, [31]). В дальнейшем мы будем обозначать константы диссоциации активатора и репрессора как K A или K R , так что (3) при полной кооперативности, в то время как (4) если происходит частичная кооперативность для TF.Здесь (⋅) + обозначает функцию положительной части (( x ) + = x , если x > 0, и ( x ) + = 0, если x ≤ 0) . Это необходимо, потому что кооперативность не будет иметь место, если в конфигурации не присутствуют два или более кооперативных TF. В оставшейся части статьи мы будем обозначать случаи, когда TF взаимодействуют между собой полностью или частично, как {{ A , R } c } и соответственно.Обратите внимание, что это обозначение охватывает случай отсутствия взаимодействия, поскольку оно соответствует случаю {{ A , R } 1 } или эквивалентно {{ A } 1 , { R } 1 }.
Места связывания упорядочены пространственно, и, как правило, нет уникального пространственного распределения для конфигурации с j A активаторами, j R репрессорами и n — j A — j R бесплатных сайтов.Например, если мы рассмотрим j A = j R = 1, есть шесть возможных пространственных распределений с одинаковыми элементами ( ARO , RAO , AOR , ROA , OAR , ORA , где O обозначает пустое пространство). В нашем описании пространственная локализация связанных частиц не рассматривается. Фактически, для конкретной конфигурации с j A активаторами, j R репрессорами и j 0 свободными сайтами возможны различные пространственные конфигурации, где k ! обозначает факториал k .
Что касается процесса связывания РНК-полимеразы промотора, TF работают вместе, пытаясь стимулировать или репрессировать процесс связывания [8] с помощью механизма, известного как рекрутинг [12, 13]. Таким образом, мы считаем, что активаторы взаимодействуют с РНКП «адгезивным» взаимодействием [22], которое приводит к модификации аффинности связывания РНК-полимеразы: где a — константа кооперативности больше 1. Напротив, эффект j R репрессоров моделируется с точки зрения «отталкивающего» взаимодействия, которое изменяет аффинность связывания с фактором антикооперативности r <1 (репрессорное взаимодействие).Мы будем называть эти параметры интенсивностью активации / репрессии транскрипции TF.
Используя предыдущие рекомендации, мы теперь опишем концентрации всех возможных конфигураций, как это было сделано в [15, 16]:
Шаг 2: Определение вероятности.
После того, как мы описали все возможные конфигурации с точки зрения концентраций активатора, репрессора и РНК-полимеразы, мы легко получаем вероятность обнаружения промотора в конкретной конфигурации j P РНК-полимеразы и j A , j R TF, связанные отношением кооперативности как (8) для всех ( j A , j R , j P ) ∈ Ω.
Шаг 3: Определение оператора BEWARE.
На этом последнем шаге оператор BEWARE получается в терминах вероятностей P ( n ) . Следуя работе Shea et al [16], синтез определенного белка зависит от общей вероятности обнаружения РНК-полимеразы в промоторе, в частности, синтез пропорционален маргинальному распределению случая j P = 1 [10, 22, 23]. Это оправдывает определение оператора BEWARE как где в определении (8) предполагается выражение (5), а C B — константа пропорциональности, которая может зависеть от других факторов, не рассмотренных в данной работе.Разделив знаменатель на две суммы, когда РНК-полимераза связана или не связана с конфигурацией, это выражение можно переписать в терминах функции фактора регуляции, F reg : (9)
Выполняя базовую алгебру, этот фактор регулирования можно уменьшить, чтобы облегчить понимание общего процесса (см. Раздел A в файле S1). Это было сделано с помощью классической стратегии, применявшейся для получения общего уравнения связывания более века назад [32].В данном контексте, насколько известно авторам, это еще не применялось. Фактически, мы можем доказать, что коэффициент регулирования может быть эквивалентно записан как (10) где явное выражение зависит от предполагаемого вида кооперативности, т. е. (11) (12) (13) для случаев некооперативного, полного и частичного сотрудничества соответственно. Обратите внимание, что математическая сложность этих выражений в основном связана с предполагаемой кооперативностью.
Варианты логики транскрипции при наличии противоположных градиентов
В этом разделе мы собираемся описать, какой будет транскрипционная реакция генов, контролируемых одними и теми же противостоящими TF, когда между ними существуют биохимические различия.Как мы объяснили в разделе «Результаты», последствия таких различий будут зависеть от типа кооперативности, происходящей между TF. Анализ случая одиночных градиентов можно найти в разделе K файла S1.
Транскрипционная логика в случае противоположных градиентов и отсутствия / полной кооперативности между TF.
Мы видим, что выражение (12) совпадает с (11), когда c = 1, что позволяет нам использовать одно и то же математическое выражение для обоих случаев, некооперативности и полной кооперативности.Итак, в обоих случаях скорости транскрипции даются формулой (9) с (14) является фактором регулирования, где c = 1, если нет кооперативности между TF, и c > 1 общая кооперативность. Фактически, мы доказываем, что логика транскрипции будет в основном одинаковой в обоих случаях.
Определение чистых концентраций активации / репрессии и диапазонов активации клеток.
Благодаря оператору BEWARE мы можем теоретически описать, какие концентрации активаторов и репрессоров вызовут более высокую или более низкую экспрессию генов, чем базальный уровень, то есть мы можем очень подробно описать эффект баланса обоих сигналов.Обратите внимание, что базальное состояние, определяемое отсутствием TF, то есть [ A ] = [ R ] = 0, соответствует F reg = 1 в выражении (9). Таким образом, фактор регуляции описывает эффективное увеличение (для F reg > 1) или уменьшение (для F reg <1) количества молекул РНКП, связанных с промотором, относительно до базального уровня, как установлено в [22].
Мы видим, что в случае (14) порог между концентрациями активации / репрессии (то есть F reg = 1) определяется линейным соотношением (15) разделив плоскость [ A ] — [ R ] на две части, мы можем обозначить область активации, если и область репрессии, если наоборот. См. Панель A на рис. S1, где показаны примеры этих пороговых значений для различных значений параметров. Очевидно, что порог (15) является линейной зависимостью между концентрациями активаторов и репрессоров.Крутизна этой прямой: не зависит от значений c и n , зависит от сродства ТФ через соотношение K R / K A , увеличивается по отношению к до a и уменьшается относительно r (так как r <1). Это оправдывает поведение чистых активированных клеточных диапазонов в случае не / полной кооперативности, указанной в таблице 1, как мы сейчас объясним.
Мы можем воспользоваться этой информацией и, рассматривая соответствующие градиенты активаторов и репрессоров, мы можем определить тканевые области чистых активированных и чистых репрессированных клеток, то есть областей клеток, экспрессирующих больше или меньше базального уровня экспрессии (см. Рис. детальное объяснение). Для ясности и принимая во внимание, что наша основная цель — понять, как эти механизмы могут изменять экспрессию генов-мишеней Hh, мы собираемся предположить, что факторы транскрипции действуют так же, как Cubitus работает в дрозофиле . Система .Hh, секретируемый из заднего отдела в передний отсек имагинального диска крыла, приводит к противоположным градиентам активатора и репрессора Ci. Граница A / P расположена примерно на 60% дорсо-вентральной (D / V) оси. Влияние градиента Hh можно оценить примерно в середине переднего компартмента, в частности, в клетках, расположенных в области между 30% и 60% по оси D / V, приблизительно. Чтобы смоделировать эти распределения концентраций TF, мы будем предполагать, что они не меняются со временем и оба являются монотонными вдоль ткани, (16) в единицах x , расстояние от границы A / P.См. Пример на вставке на панели B на рис. 1, где [CiA] / [CiR] уменьшается / увеличивается от границы A / P. Обоснуем предполагаемый ранее монотонный характер градиентов ТФ в развитии Drosophila . Хорошо известно, что концентрации CiA зависят от градиента Hh, секретируемого из заднего компартмента. Следовательно, мы будем считать, что эта концентрация уменьшается с удалением от границы A / P, то есть [ A ] ( x ) уменьшается. С другой стороны, уровни Ci-транскриптов высоки в положениях компартмента A, дистальнее границы A / P, тогда как уровни низки в положениях, проксимальных к границе A / P.Предполагая пропорциональность между уровнями транскрипции и концентрациями белка, это будет означать, что общее количество Ci (17) — неубывающая функция по отношению к расстоянию от границы A / P, x . Следовательно, концентрация CiR должна увеличиваться с удалением от границы A / P. Частным случаем такой ситуации является сохранение общего количества ТФ, предложенное в [10], соответствующее (18) равная ч , общая концентрация TF и D представляет собой крутизну градиента.Для простоты мы будем использовать гипотезу (18) на всех рисунках этой работы, хотя аналитические результаты будут рассматривать более общий случай (17). В соответствии с (18) концентрации будут ограничены прямой линией в плоскости [ A ] — [ R ] (см. Панель A на рис. S1 и панели A, C и E на рис. S2 и S3). На вставках к рисункам 1–3 и S1 и S2 на рисунках показаны распределения [ A ], [ R ]. Точки пересечения между прямой линией (18) и порогами, ([ A ] -й , [ R ] -й ) (представлены черными кружками на панели A на S1 Fig. A, C и E на фигурах S2 и S3) будут определять границу между генетически активированными и репрессированными клетками.То есть репрессированные клетки — это клетки, содержащие концентрации ([ A ], [ R ]), проверяющие (18) и [ A ] <[ A ] th . Для активированных клеток это будет [ A ]> [ A ] th . Как следствие, они будут выражать скорость транскрипции ниже / выше, чем базовая. Из-за монотонного характера распределений TF (16) активированные клетки находятся ближе к границе A / P, и предел процентной доли крылового имагинального диска, занятого активированными клетками, будет определяться расстоянием x th предоставлено Этот предел представлен синими кружками на панелях B, C и D на рис. S1 и на панелях B, D и F на рис.В любом случае проверяется, что чем ниже [ A ] th , тем больше NAC. Это, (19) по монотонному характеру градиента активатора, как это видно на S3 Рис.
Транскрипционные последствия дифференциальных биохимических характеристик.
Теперь, используя предыдущие соображения, мы хотим обосновать логику транскрипции при наличии полной кооперативности или при отсутствии какой-либо кооперативности, результаты собраны в таблице 1, столбец a) и представлены на S1 рис.В этом случае довольно легко увидеть поведение сотового диапазона, активированного сетью. Уравнение (15), которое определяет порог между концентрациями активации / репрессии, не зависит от количества энхансеров n или кооперативности c и зависит от сродства TF в терминах отношения K R / К А . Таким образом, эти области не изменяются (NAC) для генов gene1 и gene2 , так что:
- Сродство к сайтам связывания gene1 пропорционально меньше, чем сродство к сайтам связывания gene2 .С точки зрения констант диссоциации, это может быть выражено как 0 < δ <1, и это происходит потому, что мы рассматриваем пропорциональное изменение сродства к активатору и репрессорам,.
- TF взаимодействуют менее интенсивно для gene1 , чем для gene2 , то есть c g 1 < c g 2 .
- gene1 имеет меньше сайтов связывания, чем gene2 , то есть n g 1 < n g 2 .
Здесь верхние индексы g 1 и g 2 обозначают параметры генов gene1 и gene2 соответственно. Однако в случае дифференциального сродства, когда пропорциональность не проверена, чистый активированный клеточный диапазон изменится. Например, если тогда чистый активированный диапазон клеток для gene1 будет уже, чем для gene2 .
Остальные утверждения в таблице 1, столбец a) требуют некоторых простых свойств монотонности, которые были проверены с помощью лемм C и D в файле S1.Было доказано, что биохимические различия, пронумерованные 1), 2) и 3) , подтверждают, что а также Обратите внимание, что оператор BEWARE монотонно увеличивается по отношению к регуляционному фактору F reg , что позволяет нам экстраполировать эти оценки на уровни экспрессии. Эти три результата были интерпретированы как ослабление сигнала. Биохимические различия 1), 2) и 3) не может изменять характер сетевой активации / репрессии, но может сделать передачу сигналов менее эффективной.То есть они не изменяют диапазон NAC, но вызывают меньшую активацию в активированной области и меньшую репрессию в репрессированной области (↓ Sig). Следовательно, мы можем сказать, что в ситуациях 1), 2) и 3) уровни экспрессии будут уменьшаться в чистом активированном клеточном диапазоне и увеличиваться в репрессированных клетках, что ослабит диапазон клеточной экспрессии. См. S1 рис., Где изображены эти вариации.
Напротив, утверждение 3) в лемме D в файле S1 означает, что
- Если A является более слабым активатором транскрипции в gene1 , чем в gene2 , проявляя более низкую кооперативность между активаторами и РНК-полимеразой ( a g 1 < a 2131 ), то экспрессия gene1 будет меньше, чем у gene2 , потому что Очевидно, это связано с более низкими скоростями транскрипции (↓ Act) и более ограниченными сетевыми активированными клеточными диапазонами (↓ NAC) для gene1 , чем для gene2 .
- Если R является более слабым репрессором транскрипции в gene1 , чем в gene2 , проявляя более низкую антикооперативность между репрессорами и РНК-полимеразой ( r g 2 < r g 32 1 <1), то скорость экспрессии gene1 будет выше, чем экспрессия gene2 , потому что Очевидно, это включает в себя глобально более высокие скорости транскрипции (↓ Rep) и более широкие чистые активированные клеточные диапазоны (↑ NAC) для gene1 , чем для gene2 .
Логика транскрипции в случае частичной кооперативности между TF.
В случае частичной кооперативности уровни экспрессии задаются формулой (9), где коэффициент регулирования определяется как: По сравнению с (14), в этом сложном представлении скоростей экспрессии порог активации не так ясен. Итак, нам нужно провести более тонкий математический анализ. Фактически, если мы наложим пороговое уравнение (20) можно показать, что если n = 3 и c A , c R ≥ 1, этот порог определяется уникальной функцией увеличения f , проверяя (21) Он разделяет концентрации [A] — [R] на те, которые вызывают сетевую активацию, когда [ R ] / K R < f ([ A ] / K A ), и те, которые вызывают чистую репрессию, когда [ R ] / K R > f ([ A ] / K A )) (см. E в файле S1 для определения и анализа функции f ).Обратите внимание, что порог оператора BEWARE с частичной кооперативностью, в общем, не является прямой линией, хотя он показывает линейную асимптотику для больших концентраций (см. Панели A, C и E на S2 и S3 рис.). Отметим также, что тот же анализ подразумевает, что функция f не зависит от сродства TF: K A и K R .
Мы можем ясно понять эффекты частичной кооперативности в определенных предельных режимах, таких как:
- Сотрудничество только между репрессорами, то есть c A = 1 и c R > 1.Было доказано, что уровни экспрессии монотонно убывают по отношению к кооперативности репрессоров, то есть чем больше кооперативность, тем меньше экспрессия, потому что кооперативность увеличивает эффективность репрессии (см. Лемму F 2) в файле S1). Тогда, очевидно, меньшая кооперативность подразумевает меньшее подавление, провоцирующее более широкие NAC и CER.
- Взаимодействие только между активаторами, то есть c A > 1 и c R = 1.Аналогичные результаты в этом другом случае показывают, что скорость экспрессии увеличивается с кооперативностью между активаторами, поскольку это увеличивает эффективность активации (см. Лемму F 1) в файле S1). Затем уменьшение взаимодействия между активаторами снижает NAC и CER.
Эти результаты суммированы в ячейках таблицы 1 2b) и 2c) и представлены на панели D на рис. S2 и S3.
Когда оба TF взаимодействуют одновременно, мы сравнили пороги, заданные функциями f , с линейным соотношением (15), получив сравнение критериев с точки зрения следующих переменных (22) (23) В зависимости от положительного или отрицательного знака этих значений можно доказать, что переход от частичной кооперативности к некооперативности вызывает:
- Если,: мы видим уменьшение в чистом активированном диапазоне клеток по сравнению с некооперативным случаем.
- Если,: мы видим прирост в чистом активированном диапазоне ячеек по сравнению с некооперативным случаем.
- Во всех остальных случаях: увеличение или уменьшение чистого активированного клеточного диапазона может происходить в зависимости от общего количества рассматриваемых TF ( h ), аффинности связывания, K A , K R , а также интенсивности активации / репрессии. Подробное объяснение можно найти в разделе E в файле S1.
Еще один интересный аспект — это то, как эти пороги зависят от сродства TF в присутствии частичной кооперативности. Давайте снова рассмотрим ген2 с сайтами связывания с высокой аффинностью и ген 1 с сайтами связывания с пропорционально низкой аффинностью, то есть (24) Рассмотрим теперь функцию f . Он определяет порог оператора BEWARE с частичной кооперативностью, определяемый константами кооперативности a , r , c A и c R и выражением (21).Эта функция и соответствующие аффинности определяют пороговые значения концентрации активатора для gene1 и gene2 , и, используя эти выражения (25) где (17) учтено. Тогда по гипотезе (24) можно определить порядок и. Можно доказать, что, когда между ними взаимодействуют только активаторы, эти предельные значения подтверждают подразумевая, что gene2 имеет более широкий диапазон экспрессии, чем gene1 из-за (19). С другой стороны, когда только репрессоры сотрудничают, обратная связь верна. подразумевая, что gene2 имеет более узкий диапазон экспрессии, чем gene1 (подробности см. в лемме J в файле S1).На панелях D и F на рис. 3 обе эти ситуации были изображены, когда частичная кооперативность проявляется либо для активаторов, либо для репрессоров. Более подробные представления о порогах и процессе определения этих диапазонов можно найти на панелях A и B на S2 и S3 рис.
Предыдущие результаты могут быть обобщены на случай, когда оба вида взаимодействуют одновременно, а монотонность связана с условиями вогнутости и выпуклости пороговой функции f .Это состояние можно понять с точки зрения эффектов «больше, чем аддитив» и «меньше, чем аддитив», описанных в активации транскрипции [23]. Вогнутость f такая же, как при наблюдении эффектов большего, чем аддитивный, на базальном уровне. То есть, когда f является вогнутым, любые выпуклые комбинации двух различных пар концентраций, [ A ], [ R ], которые дают базальную экспрессию, будут производить чистые активированные клетки. С другой стороны, когда f является выпуклым, могут наблюдаться менее аддитивные эффекты.Этот результат и наша интерпретация контрастируют с анализом в [11] для случая кооперативных репрессоров.
Утверждения в таблице 1, ячейки 4b) и 5c) могут быть легко проверены из утверждения 3) Лемма F в файле S1.
Благодарности
O. S. благодарит профессора J. García-Ojalvo за указание ссылки [22] и R. Bielawski за редактирование и корректуру. Авторы также хотели бы поблагодарить рецензентов за их вклад.
Список литературы
- 1. Табата Т., Такей Ю. Морфогены, их идентификация и регуляция. Разработка. 2004. 131: 703–712. pmid: 14757636
- 2. Torroja C, Gorfinkiel N, Guerrero I. Механизмы формирования и интерпретации градиента ежа. Dev Neurobiol. 2005. 64: 334–356.
- 3. Халликас О., Палин К., Синюшина Н., Раутиайнен Р., Партанен Дж., Укконен Э. и др. Полногеномное прогнозирование энхансеров млекопитающих на основе анализа сродства связывания транскрипционного фактора, Cell.2006. 124 (1): 47–59. pmid: 16413481
- 4. Лорбербаум Д.С., Рамос А.И., Петерсон К.А., Карпентер Б.С., Паркер Д.С., Де С. и др. Древняя, но гибкая архитектура цис-регулирования позволяет локализовать настройку Hedgehog с помощью исправленного / Ptch2. eLife. 2016; 5: e13550. pmid: 27146892
- 5. Рамос А.И., Бароло С. Сайты связывания факторов транскрипции с низким сродством формируют ответы морфогенов и эволюцию энхансеров. Филос Т. Р. Соц Лон Б. 2013; 368 (1632): 20130017.
- 6. Gurdziel K, Lorberbaum DS, Udager AM, Song JY, Richards N, Parker DS и др.Идентификация и проверка новых реагирующих на Hedgehog энхансеров, предсказанных с помощью компьютерного анализа плотности сайтов связывания Ci / Gli. PLoS ONE. 2015; 10 (12): e0145225. pmid: 26710299
- 7. Александр С, Хасинто А, Ингам П.В. Активация транскрипции генов-мишеней hedgehog у Drosophila опосредуется непосредственно белком cubitus interruptus, членом семейства GLI ДНК-связывающих белков цинкового пальца. Джин Дев. 1996; 10: 2003–2013. pmid: 8769644
- 8. Мюллер Б., Баслер К.Репрессорные и активаторные формы Cubitus interruptus контролируют гены-мишени Hedgehog через общие общие сайты связывания Gli. Разработка. 2000; 127: 2999–3007. pmid: 10862738
- 9. Агилар-Идальго Д., Бесерра-Алонсо Д., Гарсиа-Моралес Д., Касарес Ф. К изучению ограничений регуляции генов для морфологической эволюции глазковой области дрозофилы. Dev Genes Evol. 2016; 226: 221–233. pmid: 27038024
- 10. Паркер Д.С., Уайт М.А., Рамос А.И., Коэн Б.А., Бароло С.Цис-регуляторная логика градиентных ответов ежа: ключевые роли для сродства связывания Gli, конкуренции и кооперативности. Sci Signal. 2011; 4: 1–16.
- 11. Уайт М.А., Паркер Д.С., Бароло С., Коэн Б.А. Модель пространственно ограниченной транскрипции в противоположных градиентах активаторов и репрессоров. Mol Syst Biol. 2012; 8: 614.
- 12. Пташне М., Ганн А. Активация транскреции путем набора персонала. Природа. 1997. 386 (10): 569–577. pmid: 80
- 13.Пташне М. Регуляция транскрипции: от лямбда к эукариотам. Trends Biochem Sci. 2005. 30 (6): 275–279. pmid: 15950866
- 14. Piepenburg O, Vorbrüggen G, Jäckle H. Границы сегментов дрозофилы возникают в результате одностороннего подавления активности hedgehog с помощью бескрылых сигналов. Mol Cell. 2000. 6: 203–209. pmid: 10949042
- 15. Акерс Г.К., Джонсон А.Д., Ши М.А. Количественная модель регуляции генов с помощью репрессора фага λ. P Natl Acad Sci USA, 1982; 79: 1129–1133.
- 16. Ши М., Акерс Г.К. O R Система управления бактериофагом лямбда. Физико-химическая модель регуляции генов. J Mol Biol. 1985; 181: 211–230. pmid: 3157005
- 17. Агилар-Идальго Д., Домингес-Сехудо М.А., Аморе Дж., Брокманн А., Лемос М.С., Кордова А. и др. Генная сеть, управляемая Hh, контролирует спецификацию, структуру и размер простых глаз дрозофилы. Разработка. 2013; 140: 82–92. pmid: 23154412
- 18.Aguilar-Hidalgo D, Lemos MC, Córdoba A. Основной мотив регуляторной сети лежит в основе формирования паттерна глазкового комплекса у Drosophila melanogaster. Физика Д. 2015; 295: 91–102.
- 19. Гилман А, Аркин А.П. Генетический «код»: представление и динамические модели генетических компонентов и сетей. Анну Рев Геном Хам Г. 2002; 3: 341–369.
- 20. Ай А, Арности Д.Н. Математическое моделирование экспрессии генов: руководство для озадаченного биолога. Crit Rev Biochem Mol.2011. 46 (2): 137–151.
- 21. Франк Т.Д., Кавадас МАС, Нгуен Л.К., Чеонг А. Нелинейная динамика в регуляции транскрипции: биологические логические ворота. В: Carballido-Landeira J, Escribano B, редакторы. Нелинейная динамика в биологических системах, серия SEMA SIMAI Springer. 2016; 7: 43–62.
- 22. Bintu L, Buchler NE, García HG, Gerland U, Hwa T., Kondev J, et al. Регуляция транскрипции в цифрах: модели. Curr Opin Genet Dev. 2005. 15: 116–124. pmid: 15797194
- 23.Франк Т.Д., Кармоди А.М., Холоденко Б.Н. Универсальность совместной активации транскрипции: анализ термодинамического моделирования для более сильных и менее аддитивных эффектов. PLoS ONE. 2012; 7 (4): e34439. pmid: 22506020
- 24. Zhou X, Su Z. tCal: калькулятор транскрипционной вероятности с использованием термодинамической модели. Биоинформатика. 2008. 24 (22): 2639–2640. pmid: 18796477
- 25. Роджерс К.В., Шир А.Ф. Градиенты морфогенов: от поколения к интерпретации, Annu Rev Cell Dev Bi.2011; 27: 377–407.
- 26. Юнкер Дж. П., Петерсон К. А., Ниши Ю., Мао Дж., МакМахон А. П., ван Ауденаарден А. Прогностическая модель передачи сигналов бифункционального транскрипционного фактора во время формирования эмбрионального тканевого паттерна. Dev Cell. 2014; 31: 448–460. pmid: 25458012
- 27. Balaskas N, Ribeiro A, Panovska J, Dessaud E, Sasai N, Page KM, et al. Генетическая регуляторная логика для чтения градиента передачи сигналов Sonic Hedgehog в нервной трубке позвоночных. Клетка. 2012; 148: 273–284 pmid: 22265416
- 28.Кичева А., Болленбах Т., Рибейро А., Валле Х.П., Ловелл-Бэдж Р, Эпископоу В. и др. Координация спецификации и роста предков в спинном мозге мышей и кур. Наука. 2014; 345: 1254927. pmid: 25258086
- 29. Саха К., Шаффер Д.В. Динамика сигнала в формировании паттерна ткани Sonic hedgehog. Разработка. 2006. 133: 889–900. pmid: 16452094
- 30. Вербени М., Санчес О., Моллика Э., Зигль-Кашеденье И., Карлентон А., Герреро И. и др. Морфогенетическое действие за счет распространения, ограниченного потоком.Phys Life Rev.2013; 10: 457–475. pmid: 23831049
- 31. Лай К., Робертсон М.Дж., Шаффер Д.В. Сигнальная система Sonic Hedgehog как бистабильный генетический переключатель. Биофиз Дж. 2004; 86: 2748–2757. pmid: 15111393
- 32. Биссвангер Х. Ферментная кинетика: принципы и методы. 2-е изд. Вайнхайм: ВИЛИ-ВЧ; 2008.
Наследственный гемохроматоз (HH) — Американский колледж гастроэнтерологии
Лечение простое, недорогое и безопасное.Первый шаг — избавить организм от лишнего железа. Этот процесс называется флеботомией, что означает удаление крови так же, как она берется у доноров в банках крови. В зависимости от тяжести перегрузки железом один или два раза в неделю берут пол-литра крови (500 мл) в течение от нескольких месяцев до года, а иногда и дольше. Кровопускание одной единицы крови обычно снижает уровень ферритина примерно на 30 мкг / л. Уровень ферритина в крови периодически проверяется для контроля уровня железа. Цель состоит в том, чтобы довести уровень ферритина в крови до нижнего предела нормы и удерживать его на этом уровне.Это означает снижение содержания ферритина до 50-100 нг / л.
Когда уровень железа возвращается к норме, начинается поддерживающая терапия, которая включает удаление пол-литра крови каждые 2–4 месяца на всю жизнь. Некоторым людям может потребоваться более частая флеботомия. Многие пациенты, которым проводят поддерживающую флеботомию, делают это как доноры крови. Ежегодный анализ крови на ферритин поможет определить, как часто следует удалять кровь. Также необходимо регулярное наблюдение у специалиста, который интересуется гемохроматозом или расстройством, связанным с перегрузкой железом.Это может быть гематолог (врач крови) или гастроэнтеролог / гепатолог (пищеварительный или печеночный врач).
Если лечение начинается до того, как органы будут повреждены, можно предотвратить сопутствующие заболевания, такие как заболевания печени, сердца, артрит и диабет. Перспективы людей, у которых на момент постановки диагноза уже были эти состояния, зависят от степени поражения органа. Например, лечение HH может остановить прогрессирование заболевания печени на ранних стадиях, что приводит к нормальной продолжительности жизни.Однако, если развился цирроз или рубцевание печени, риск развития рака печени увеличивается, даже если запасы железа уменьшаются до нормального уровня. Люди с диабетом, возникшим в результате повреждения поджелудочной железы, обычно видят улучшение, если не обращение вспять, в зависимости от того, сколько повреждений произошло. Лечение не может вылечить состояния, связанные с установленным ДГ, но поможет улучшить большинство из них. Аномальные тесты печени, сердечная дисфункция и утомляемость могут улучшиться при лечении, но боль в суставах, диабет и снижение полового влечения — нет.Пигментация кожи после лечения постепенно улучшается.
При проведении флеботомии нет необходимости ограничивать количество железа в пище. Людям с HH не следует принимать добавки с железом или витамином C. Пациентам с поражением печени не следует употреблять алкогольные напитки, потому что они могут еще больше повредить печень, или есть сырые морепродукты из-за риска серьезной инфекции.
Скрининг на ДГ — тестирование людей, у которых нет симптомов, — не является рутинной частью медицинского обслуживания или обследований.Тем не менее, братья и сестры пациентов с ГП должны сдать кровь на анализ, чтобы узнать, есть ли у них заболевание или являются ли они носителями. Родственникам первой степени родства пациентов с этим заболеванием следует предложить пройти обследование. Тестирование должно включать анализ насыщения трансферрина, ферритина и мутации
HFE (генный тест). При рассмотрении возможности тестирования детей пациента может быть проще сначала проверить другого родителя. Если он / она отрицателен в отношении каких-либо мутацийHFE
, то дети могут быть только носителями.Однако, если у другого родителя есть генная мутация, детям следует предложить генетическое тестирование. Каждый раз, когда проводится генетическое тестирование, должен быть доступен сертифицированный генетический консультант и должно быть получено информированное согласие.Врачи должны посоветовать пациентам пройти обследование их близких родственников и рассмотреть возможность обследования людей с заболеваниями суставов, тяжелой и продолжающейся усталостью, сердечными заболеваниями, повышенными ферментами печени, импотенцией и диабетом, поскольку эти состояния могут быть результатом ГГ.
У пациентов с ГГ без цирроза печени показатели выживаемости не отличаются от показателей общей популяции, тогда как выживаемость пациентов с ГГ и циррозом печени значительно ниже.
Во время начальной, а также поддерживающей фазы лечения кровь, взятая у пациента с ДГ, использовалась для переливаний. Не существует универсальных правил в отношении того, следует ли использовать кровь от пациентов с ДГ для донорства. Многие банки крови придерживаются политики запрета приема крови от пациентов с ДГ, хотя другие сделали это без каких-либо осложнений.
Автор (ы) и дата (ы) публикации
Брюс Р.Бэкон, доктор медицины, FACG, и Судханшу Гогиа, доктор медицины, Медицинский факультет Университета Сент-Луиса, Сент-Луис, штат Миссури, — опубликовано в феврале 2011 г.
Обновлено 2020 г., Пранав Б. Патель, доктор медицины, Монтгомери, штат Алабама, для Комитета по уходу за пациентами ACG
Вернуться к началу
Анализ транскрипционной логики, управляющей дифференциальной пространственной экспрессией в генах-мишенях Hh
Выведение оператора BEWARE для двух противоположных градиентов TF
В качестве первого шага мы применяем идеи статистического термодинамического метода к гену, g , контролируется двумя противоположными факторами транскрипции { A , R }, активатором и репрессором соответственно.Наша цель здесь состоит в том, чтобы вывести выражения для изменения концентрации белка G с течением времени в терминах концентраций A и R, [ A ] и [ R ], т.е.
d [G] dt = ОСТОРОЖНО ([A], [R]).
(1)
Здесь «BEWARE ()» представляет собой математическую функцию, определяющую зависимость относительно роли активации / репрессии TF. Это не зависит от других возможных факторов, имеющих отношение к эволюции белка, например, деградации или пространственной дисперсии.В модели реакции связывания TF и RNAP в энхансерах и промоторе, соответственно, намного быстрее, чем синтез белка G, поэтому он будет рассматриваться в термодинамическом равновесии, определяемом Законом действия масс. Если B представляет собой пустую регуляторную область, набор незанятых энхансеров-промотор, комплексы BA , BR и BRNAP имеют концентрацию в равновесии, определяемую
[BA] = k + A (1) kA (1) [A] [B]: = [A] KA (1) [B],
[BR] = k + R (1) kR (1 ) [R] [B]: = [R] KR (1) [B],
[BRNAP] = k + RPk-RP [RNAP] [B]: = [RNAP] KRP [B],
где KA (1), KR (1), K RP , [ A ], [ R ] и [ RNAP ] — константы диссоциации и концентрации активаторов, репрессоров и РНК-полимеразы.Таким образом, отношения [A] KA (1), [R] KR (1) и [RNAP] KRP безразмерны. Верхний индекс (1) обозначает константу диссоциации реакции, которая происходит в отсутствие другого TF, ранее связанного с другим энхансером (обратите внимание, что, поскольку наборы имеют только один промотор, верхний индекс не нужен для константы диссоциации RNAP) . Заметим, что чем выше сродство между белком / комплексом и сайтом связывания, тем ниже константа диссоциации в соответствующей реакции связывания.Последовательное связывание более одного фактора транскрипции считается последовательным и конкурентным процессом, так что реакции
A + BA⇌k − A (2) k + A (2) BAAorR + BA⇌k − R (2) k + R (2) BAR
задаются равновесными концентрациями
[BAA] = [A] [A] KA (1) KA (2) [B] и [BAR] = [A] [R] KA (1) KR (2) [B] »,
здесь верхний индекс (2) обозначает константу диссоциации для реакции TF, который уже связывает оператора с одним TF в другом сайте. Конкуренция моделируется таким образом, что константа диссоциации конфигурации свободных сайтов не зависит от их положения, но может зависеть от других TF, уже связанных с тем же набором энхансеров посредством кооперативности или антикооперативности.
Мы обозначаем как некооперативные ТФ все те белки, сродство к энхансеру которых не модифицируется никакими ранее связанными ТФ, то есть они подтверждают, что KA (2) = KA (1) и KR (2) = KR (1). Это предположение подразумевает последовательную независимость равновесных концентраций, поскольку [ BRA ] = [ BAR ]. Вероятно предположить то же соотношение для последующих привязок, то есть KA (j) = KA (1) и KR (j) = KR (1) для j ≥ 2. Вследствие этой последовательной независимости мы обозначаем константы диссоциации равны K A и K R без надстрочного индекса.Таким образом, если все рассматриваемые TF не кооперативны, мы легко делаем вывод, что концентрация в равновесии конфигурации с j A активаторами и j R связанными репрессорами, равна
[BAjARjR] = [B] ([A] KA) jA ([R] KR) jR
(2)
независимо от последовательного порядка связывания и конкретных позиций, занимаемых TF. Хотя цис-регуляторные элементы дикого типа Drosophila включают в себя всего 3 сайта связывания, мы собираемся сравнить с экспериментами, в которых эти сайты связывания были уменьшены до 1.Таким образом, мы будем рассматривать в нашей модели n ≥ 1 количество сайтов связывания ТФ. Во всех случаях у нас есть ограничение на возможное количество связанных факторов транскрипции. Итак, j A + j R ≤ n должен быть проверен, и, как следствие, j 0 = n — j A — j R ≥ 0 обозначает количество свободных мест в конфигурации.
С другой стороны, кооперативность возникает, когда существование других ранее связанных белков влияет на сродство нового связывающего белка типа i , k = A , R , то есть:
где c — положительная константа, больше 1, если белки взаимодействуют, и меньше 1, если происходит антикооперативность. Поскольку единственная разница между кооперативностью и антикооперативностью — это пороговое значение для c , в последующем моделировании мы будем ссылаться на константу c и не будем различать оба случая.Если кооперативность имеет место, необходимо знать, на какие TF влияют другие TF, поскольку равновесная концентрация будет зависеть от этих соотношений. В предыдущей литературе недавно было предложено, чтобы полная и частичная кооперативность играла важную роль в генах-мишенях Hh / Shh посредством Ci / Gli TFs [10, 11, 26]. Частичная кооперативность активаторов будет происходить, когда наличие связанного активатора одинаково изменяет аффинность связывания любого заднего активатора, то есть KA (j) = KA (1) / cA для j ≥ 2.То же самое и с репрессорами. Полная кооперативность будет иметь место, когда присутствие связанного TF изменяет аффинность любого заднего связывания таким же образом, то есть KA (j) = KA (1) / c и одновременно KR (j) = KR (1) / c для j ≥ 2 (см., например, [31]). В дальнейшем мы будем обозначать константы диссоциации активатора и репрессора как K A или K R , так что
[BAjARjR] = [B] c (jA + jR-1) + ([A] KA) jA ([R] KR) jR
(3)
при полной кооперативности, в то время как
[BAjARjR] = [B] cA (jA-1) + cR (jR-1) + ([A] KA) jA ([R] KR) jR
(4)
если происходит частичная кооперативность для TF.Здесь (⋅) + обозначает функцию положительной части (( x ) + = x , если x > 0, и ( x ) + = 0, если x ≤ 0) . Это необходимо, потому что кооперативность не будет иметь место, если в конфигурации не присутствуют два или более кооперативных TF. В оставшейся части статьи мы будем обозначать случаи, когда TF взаимодействуют между собой полностью или частично, как {{ A , R } c } и {{A} cA, {R} cR} соответственно.Обратите внимание, что это обозначение охватывает случай отсутствия взаимодействия, поскольку оно соответствует случаю {{ A , R } 1 } или эквивалентно {{ A } 1 , { R } 1 }.
Места связывания упорядочены пространственно, и, как правило, нет уникального пространственного распределения для конфигурации с j A активаторами, j R репрессорами и n — j A — j R бесплатных сайтов.Например, если мы рассмотрим j A = j R = 1, есть шесть возможных пространственных распределений с одинаковыми элементами ( ARO , RAO , AOR , ROA , OAR , ORA , где O обозначает пустое пространство). В нашем описании пространственная локализация связанных частиц не рассматривается. Фактически, для конкретной конфигурации с j A активаторами, j R репрессорами и j 0 бесплатных сайтов n! J0! JA! JR! возможны разные пространственные конфигурации, где k ! обозначает факториал k .
Что касается процесса связывания РНК-полимеразы промотора, TF работают вместе, пытаясь стимулировать или репрессировать процесс связывания [8] с помощью механизма, известного как рекрутинг [12, 13]. Таким образом, мы считаем, что активаторы взаимодействуют с РНКП посредством «адгезионного» взаимодействия [22], которое приводит к модификации аффинности связывания РНК-полимеразы: KRP / ajA, где a — константа кооперативности больше 1. Напротив, Эффект репрессоров j R моделируется в терминах «отталкивающего» взаимодействия, которое изменяет аффинность связывания KRP / rjR с фактором антикооперативности r <1 (репрессорное взаимодействие).Мы будем называть эти параметры интенсивностью активации / репрессии транскрипции TF.
Используя предыдущие рекомендации, мы теперь опишем концентрации всех возможных конфигураций, как это было сделано в [15, 16]:
Шаг 1: Построение пространства образца
Все возможные способы получения равновесной концентрации с j A , j R и j P активаторы, репрессоры и РНК-полимеразы задаются состояниями
Z (n) (jA, jR, jP = 1; C) = C (C) n! J0! JA! JR! [B] [RNAP] KRP (a [A] KA) jA (r [R] KR) jR,
(5)
Z (n) (jA, jR, jP = 0; C) = C (C) n! J0! JA! JR! [B] ([A] KA) jA ( [R] KR) jR
где j P = 1 означает, что существует связанная РНК-полимераза, а j P = 0 нет, j 0 = n — j A — j R ≥ 0, а переменная C описывает отношение кооперативности между TF.В частности, используя (3) и (4), функция кооперативности C принимает значения
C (C = {A, R} c) = c (jA + jR-1) +
(6)
а также
C (C = {{A} cA, {R} cR}) = cA (jA-1) + cR (jR-1) +.
(7)
Это позволяет нам описать все пространство выборки, то есть пространство всех возможных конфигураций, с помощью
Ом = {(jA, jR, jP); jA, jR≥0, jA + jR≤n, jP = 0,1}.
Шаг 2: Определение вероятности
После того, как мы описали все возможные конфигурации с точки зрения концентраций активатора, репрессора и РНК-полимеразы, мы легко получаем вероятность обнаружения промотора в конкретной конфигурации j P РНК-полимераза и j A , j R TF, связанные отношением кооперативности C как
P (n) (jA, jR, jP; C) = Z (n) (jA, jR, jP; C) ∑ {jA ′, jR ′, jP ′} ∈ΩZ (n) (jA ′, jR ′, JP ′; В),
(8)
для всех ( j A , j R , j P ) ∈ Ω.
Шаг 3: Определение оператора BEWARE
На этом последнем шаге оператор BEWARE получается в терминах вероятностей P ( n ) . Следуя работе Shea et al [16], синтез определенного белка зависит от общей вероятности обнаружения РНК-полимеразы в промоторе, в частности, синтез пропорционален маргинальному распределению случая j P = 1 [10, 22, 23]. Это оправдывает определение оператора BEWARE как
ОСТОРОЖНО ([A], [R], [RNAP]; C) = CB∑jA, jR≥0jA + jR≤nP (n) (jA, jR, jP = 1; C)
где в определении (8) предполагается выражение (5), а C B — константа пропорциональности, которая может зависеть от других факторов, не рассмотренных в данной работе.Разделив знаменатель на две суммы, когда РНК-полимераза связана или не связана с конфигурацией, это выражение можно переписать в терминах функции фактора регуляции, F reg :
ОСТОРОЖНО ([A], [R], [RNAP]; C) = CB1 + ∑jA ′, jR′≥0jA ′ + jR′≤nZ (n) (jA ′, jR ′, jP ′ = 0; C ) ∑jA ′, jR′≥0jA ′ + jR′≤nZ (n) (jA ′, jR ′, jP ′ = 1; C) = CB1 + KRP [RNAP] Freg ([A], [R]; C ).
(9)
Выполняя базовую алгебру, этот коэффициент регулирования можно уменьшить, чтобы облегчить понимание общего процесса (см. Раздел A в файле S1).Это было сделано с помощью классической стратегии, применявшейся для получения общего уравнения связывания более века назад [32]. В данном контексте, насколько известно авторам, это еще не применялось. Фактически, мы можем доказать, что коэффициент регулирования может быть эквивалентно записан как
Freg ([A], [R]; C) = S (n) (a [A] KA-1, r [R] KR-1; C) S (n) ([A] KA-1, [R] KR-1; C),
(10)
где явное выражение S (n) (x, y; C) зависит от предполагаемого вида кооперативности, т. е.
S (n) (x, y; {{A, R} 1}) = (1 + x + y) n,
(11)
S (n) (x, y; {{A, R} c}) = 1-1c + 1c (1 + cx + cy) n,
(12)
S (n) (x, y; {{A} cA, {R} cR}) = ( 1 + cAx + cRy) ncAcR + (1-1cR) (1 + cAx) ncA + (1-1cA) (1 + cRy) ncR + (1-1cA) (1-1cR),
(13)
для случаев некооперативного, полного и частичного сотрудничества соответственно.Обратите внимание, что математическая сложность этих выражений в основном связана с предполагаемой кооперативностью.
Варианты транскрипционной логики в присутствии противоположных градиентов
В этом разделе мы собираемся описать, какой будет транскрипционная реакция генов, контролируемых одними и теми же противоположными ТФ, при наличии биохимических различий между ними. Как мы объяснили в разделе «Результаты», последствия таких различий будут зависеть от типа кооперативности, происходящей между TF.Анализ случая одиночных градиентов можно найти в разделе K файла S1.
Транскрипционная логика в случае противоположных градиентов и не / полной кооперативности между TF
Мы видим, что выражение (12) совпадает с (11), когда c = 1, что позволяет нам использовать одно и то же математическое выражение для обоих случаев, отказ от сотрудничества и полное сотрудничество. Итак, в обоих случаях скорости транскрипции даются формулой (9) с
Freg ([A], [R]; {A, R} c) = 1-1c + 1c (1 + ac [A] KA + rc [R] KR) n1-1c + 1c (1 + c [ A] KA + c [R] KR) n
(14)
является фактором регулирования, где c = 1, если нет кооперативности между TF, и c > 1 общая кооперативность.Фактически, мы доказываем, что логика транскрипции будет в основном одинаковой в обоих случаях.
Определение чистых концентраций активации / репрессии и диапазонов активации клеток
Благодаря оператору BEWARE мы можем теоретически описать, какие концентрации активаторов и репрессоров вызовут более высокую или более низкую экспрессию генов, чем базальный уровень, т.е. детализировать эффект баланса обоих сигналов. Обратите внимание, что базальное состояние, определяемое отсутствием TF, то есть [ A ] = [ R ] = 0, соответствует F reg = 1 в выражении (9).Таким образом, фактор регуляции описывает эффективное увеличение (для F reg > 1) или уменьшение (для F reg <1) количества молекул РНКП, связанных с промотором, относительно до базального уровня, как установлено в [22].
Мы видим, что в случае (14) порог между концентрациями активации / репрессии (то есть F reg = 1) определяется линейной зависимостью
разделив плоскость [ A ] — [ R ] на две части, мы можем обозначить область активации, если [R]
Мы можем взять эту информацию и, рассматривая соответствующие градиенты активаторов и репрессоров, мы можем определить тканевые области чистых активированных и чистых репрессированных клеток, то есть областей клеток, экспрессирующих больше или меньше базального уровня экспрессии (см. объяснение). Для ясности и принимая во внимание, что наша основная цель — понять, как эти механизмы могут изменять экспрессию генов-мишеней Hh, мы собираемся предположить, что факторы транскрипции действуют так же, как Cubitus работает в дрозофиле . Система .Hh, секретируемый из заднего отдела в передний отсек имагинального диска крыла, приводит к противоположным градиентам активатора и репрессора Ci. Граница A / P расположена примерно на 60% дорсо-вентральной (D / V) оси. Влияние градиента Hh можно оценить примерно в середине переднего компартмента, в частности, в клетках, расположенных в области между 30% и 60% по оси D / V, приблизительно. Чтобы смоделировать эти распределения концентраций TF, мы будем предполагать, что они не меняются со временем и оба являются монотонными вдоль ткани,
[A] = [A] (x) строго убывающий и [R] = [R] (x) строго возрастающий
(16)
в единицах x , расстояние от границы A / P.См. Пример на вставке на панели B, где [CiA] / [CiR] уменьшается / увеличивается от границы A / P. Обоснуем предполагаемый ранее монотонный характер градиентов ТФ в развитии Drosophila . Хорошо известно, что концентрации CiA зависят от градиента Hh, секретируемого из заднего компартмента. Следовательно, мы будем считать, что эта концентрация уменьшается с удалением от границы A / P, то есть [ A ] ( x ) уменьшается. С другой стороны, уровни Ci-транскриптов высоки в положениях компартмента A, дистальнее границы A / P, тогда как уровни низки в положениях, проксимальных к границе A / P.Предполагая пропорциональность между уровнями транскрипции и концентрациями белка, это будет означать, что общее количество Ci
— неубывающая функция по отношению к расстоянию от границы A / P, x . Следовательно, концентрация CiR должна увеличиваться с удалением от границы A / P. Частным случаем такой ситуации является сохранение общего количества ТФ, предложенное в [10], соответствующее
[A] = he-x / D, [R] = h- [A],
(18)
равная ч , общая концентрация TF и D представляет собой крутизну градиента.Для простоты мы будем использовать гипотезу (18) на всех рисунках этой работы, хотя аналитические результаты будут рассматривать более общий случай (17). В соответствии с (18) концентрации будут ограничены прямой линией в плоскости [ A ] — [ R ] (см. Панель A на рис. S1 и панели A, C и E на рис. S2 и S3). На вставках к фиг. — и S1 и S2 фиг. Показаны распределения [ A ], [ R ]. Точки пересечения между прямой линией (18) и порогами, ([ A ] -й , [ R ] -й ) (представлены черными кружками на панели A на S1 Fig. A, C и E на фигурах S2 и S3) будут определять границу между генетически активированными и репрессированными клетками.То есть репрессированные клетки — это клетки, содержащие концентрации ([ A ], [ R ]), проверяющие (18) и [ A ] <[ A ] th . Для активированных клеток это будет [ A ]> [ A ] th . Как следствие, они будут выражать скорость транскрипции ниже / выше, чем базовая. Из-за монотонного характера распределений TF (16) активированные клетки находятся ближе к границе A / P, и предел процентной доли крылового имагинального диска, занятого активированными клетками, будет определяться расстоянием x th предоставлено
Этот предел представлен синими кружками на панелях B, C и D на рис. S1 и на панелях B, D и F на рис.В любом случае проверяется, что чем ниже [ A ] th , тем больше NAC. Это,
[A] th2 <[A] th3 → xth2> xth3
(19)
по монотонному характеру градиента активатора, как это видно на S3 Рис.
Транскрипционные последствия дифференциальных биохимических характеристик
Теперь, используя предыдущие соображения, мы хотим обосновать логику транскрипции при наличии полной кооперативности или при отсутствии какой-либо кооперативность, результаты собраны в столбце a) и представлены в S1 Рис.В этом случае довольно легко увидеть поведение сотового диапазона, активированного сетью. Уравнение (15), которое определяет порог между концентрациями активации / репрессии, не зависит от количества энхансеров n или кооперативности c и зависит от сродства TF в терминах отношения K R / К А . Таким образом, эти области не изменяются (NAC) для генов gene1 и gene2 , так что:
Сродство к сайтам связывания gene1 меньше, чем сродство к сайтам связывания gene2 в пропорционально.С точки зрения констант диссоциации это может быть выражено как KRg2 = δKRg1 и KAg2 = δKAg1, равным 0 < δ <1, и это происходит потому, что мы рассматриваем пропорциональное изменение сродства к активатору и репрессорам, KRg1 / KAg1 = KRg2 / KAg2. .
ТФ взаимодействуют менее интенсивно для gene1 , чем для gene2 , то есть c g 1 < c g 2 .
gene1 имеет меньше сайтов связывания, чем gene2 , то есть n g 1 < n g 2 .
Здесь верхние индексы g 1 и g 2 обозначают параметры генов gene1 и gene2 соответственно. Однако в случае дифференциального сродства, когда пропорциональность не проверена, чистый активированный клеточный диапазон изменится. Например, если KRg2 / KAg2> KRg1 / KAg1, то чистый активированный клеточный диапазон для gene1 будет уже, чем для gene2 .
Остальные утверждения в столбце a) требуют некоторых простых свойств монотонности, которые были проверены с помощью лемм C и D в файле S1.Было доказано, что биохимические различия, пронумерованные 1), 2) и 3) , подтверждают, что
Fregg2> Fregg1 (> 1), если [A] и [R] относятся к области активации
а также
Fregg2 Обратите внимание, что оператор BEWARE монотонно увеличивается по отношению к регуляционному фактору F reg , что позволяет нам экстраполировать эти оценки на уровни экспрессии.Эти три результата были интерпретированы как ослабление сигнала. Биохимические различия 1), 2) и 3) не может изменять характер сетевой активации / репрессии, но может сделать передачу сигналов менее эффективной. То есть они не изменяют диапазон NAC, но вызывают меньшую активацию в активированной области и меньшую репрессию в репрессированной области (↓ Sig). Следовательно, мы можем сказать, что в ситуациях 1), 2) и 3) уровни экспрессии будут уменьшаться в чистом активированном клеточном диапазоне и увеличиваться в репрессированных клетках, что ослабит диапазон клеточной экспрессии.См. S1 рис., Где изображены эти вариации. Напротив, утверждение 3) в лемме D в файле S1 подразумевает, что Если A является более слабым активатором транскрипции в gene1 , чем в gene2 , проявляя более низкую кооперативность между активаторами и РНК. полимеразы ( a g 1 < a g 2 ), то экспрессия gene1 будет меньше, чем у gene2 , потому что Fregg1 ([A], [R]; {A, R} c) Очевидно, это связано с более низкими скоростями транскрипции (↓ Act) и более ограниченными сетевыми активированными клеточными диапазонами (↓ NAC) для gene1 , чем для gene2 . Если R является более слабым репрессором транскрипции в gene1 , чем в gene2 , проявляя более низкую антикооперативность между репрессорами и РНК-полимеразой ( r g 2 < r g 1 <1), то скорость экспрессии gene1 будет выше, чем экспрессия gene2 , потому что Fregg1 ([A], [R]; {A, R} c)> Fregg2 ([A], [R]; {A, R} c). Очевидно, это включает в себя глобально более высокие скорости транскрипции (↓ Rep) и более широкие чистые активированные клеточные диапазоны (↑ NAC) для gene1 , чем для gene2 . В случае частичной кооперативности скорости экспрессии задаются формулой (9), где фактор регуляции определяется как: Freg ([A], [R]; {{A} cA, {R} cR}) = (1 + acA [A] KA + rcR [R] KR) ncAcR + (1-1cR) (1 + cAa [A] KA) ncA + (1-1cA) (1 + cRr [R] KR) ncR + (1-1cA) (1-1cR) (1 + cA [A] KA + cR [R] KR) ncAcR + (1- 1cR) (1 + cA [A] KA) ncA + (1-1cA) (1 + cR [R] KR) ncR + (1-1cA) (1-1cR). По сравнению с (14), в этом сложном представлении скоростей экспрессии порог активации не так ясен. Итак, нам нужно провести более тонкий математический анализ. Фактически, если мы наложим пороговое уравнение Freg ([A], [R]; {{A} cA, {R} cR}) = 1, (20) можно показать, что если n = 3 и c A , c R ≥ 1, этот порог определяется уникальной функцией увеличения f , проверяя Freg ([A], KRf ([A] / KA); {{A} cA, {R} cR}) = 1. (21) Он разделяет концентрации [A] — [R] на те, которые вызывают сетевую активацию, когда [ R ] / K R < f ([ A ] / K A ), и те, которые вызывают чистую репрессию, когда [ R ] / K R > f ([ A ] / K A )) (см. E в файле S1 для определения и анализа функции f ).Обратите внимание, что порог оператора BEWARE с частичной кооперативностью, в общем, не является прямой линией, хотя он показывает линейную асимптотику для больших концентраций (см. Панели A, C и E на S2 и S3 рис.). Отметим также, что тот же анализ подразумевает, что функция f не зависит от сродства TF: K A и K R . Мы можем ясно понять эффекты частичной кооперативности в определенных предельных режимах, таких как: Кооперативность только между репрессорами, то есть c A = 1 и c R > 1 .Было доказано, что уровни экспрессии монотонно убывают по отношению к кооперативности репрессоров, то есть чем больше кооперативность, тем меньше экспрессия, потому что кооперативность увеличивает эффективность репрессии (см. Лемму F 2) в файле S1). Тогда, очевидно, меньшая кооперативность подразумевает меньшее подавление, провоцирующее более широкие NAC и CER. Взаимодействие только между активаторами, то есть c A > 1 и c R = 1.Аналогичные результаты в этом другом случае показывают, что скорость экспрессии увеличивается с кооперативностью между активаторами, поскольку это увеличивает эффективность активации (см. Лемму F 1) в файле S1). Затем уменьшение взаимодействия между активаторами снижает NAC и CER. Эти результаты суммированы в ячейках 2b) и 2c) и представлены на панели D на S2 и S3 рис. Когда оба TF взаимодействуют одновременно, мы сравнили пороги, заданные функциями f , с линейным соотношением (15), получив сравнение критериев с точки зрения следующих переменных a¯2 = 3cAcRa-1r-1 {2+ (1 + a) (r-1) cA-2ar + (a-1) (r + 1) cR}, (22) a¯3 = cAcRa-1 (r-1) 2 {(1 + a + a2) cA2 (r-1) 2-3cA (r-1) (a2r-1) — (1 + r + r2) (a-1) 2cR2 + 3 (a-1) (ar2-1) cR}. (23) В зависимости от положительного или отрицательного знака этих значений может быть доказано, что переход от частичной кооперативности к некооперативности вызывает: Если a¯2> 0, a¯3> 0: мы видим декремент в активированной сети. сотовый диапазон по отношению к некооперативному случаю. Если a¯2 <0, a¯3 <0: мы видим приращение в чистом активированном клеточном диапазоне по сравнению с некооперативным случаем. Во всех других случаях: увеличение или уменьшение чистого активированного клеточного диапазона может происходить в зависимости от общего количества рассматриваемых TF ( h ), аффинности связывания, K A , K R , а также интенсивности активации / репрессии.Подробное объяснение можно найти в разделе E в файле S1. Еще один интересный аспект — это то, как эти пороги зависят от сродства TF в присутствии частичной кооперативности. Давайте снова рассмотрим ген2 с сайтами связывания с высокой аффинностью и ген 1 с сайтами связывания с пропорционально низкой аффинностью, то есть KRg2 = δKRg1иKAg2 = δKAg1being0 <δ <1. (24) Рассмотрим теперь функцию f .Он определяет порог оператора BEWARE с частичной кооперативностью, определяемый константами кооперативности a , r , c A и c R и выражением (21). Эта функция и соответствующие аффинности определяют пороговые значения концентрации активатора для gene1 и gene2 , [A] thg1 и [A] thg2, используя эти выражения [A] thg1 + KRg1f ([A] thg1 / KAg1) = h (x) и [A] thg2 + KRg2f ([A] thg2 / KAg2) = h (x), (25) где (17) учтено.Тогда, согласно гипотезе (24), можно определить порядок [A] thg2 и [A] thg1. Можно доказать, что, когда между ними взаимодействуют только активаторы, эти предельные значения подтверждают подразумевая, что gene2 имеет более широкий диапазон экспрессии, чем gene1 из-за (19). С другой стороны, когда только репрессоры сотрудничают, обратная связь верна. подразумевая, что gene2 имеет более узкий диапазон экспрессии, чем gene1 (подробности см. в лемме J в файле S1).На панелях D и F в обеих этих ситуациях изображены случаи, когда частичная кооперативность проявляется либо для активаторов, либо для репрессоров. Более подробные представления о порогах и процессе определения этих диапазонов можно найти на панелях A и B на S2 и S3 рис. Предыдущие результаты могут быть обобщены на случай, когда оба вида взаимодействуют одновременно, а монотонность связана с условиями вогнутости и выпуклости пороговой функции f . Это состояние можно понять с точки зрения эффектов «больше, чем аддитив» и «меньше, чем аддитив», описанных в активации транскрипции [23].Вогнутость f такая же, как при наблюдении эффектов большего, чем аддитивный, на базальном уровне. То есть, когда f является вогнутым, любые выпуклые комбинации двух различных пар концентраций, [ A ], [ R ], которые дают базальную экспрессию, будут производить чистые активированные клетки. С другой стороны, когда f является выпуклым, могут наблюдаться менее аддитивные эффекты. Этот результат и наша интерпретация контрастируют с анализом в [11] для случая кооперативных репрессоров. Утверждения в ячейках 4b) и 5c) могут быть легко проверены из утверждения 3) Лемма F в файле S1. Учитывая набор объектов (входной класс объектов) и поле анализа (поле ввода), инструмент анализа кластеров и выбросов идентифицирует пространственные кластеры объектов с помощью высокие или низкие значения. Инструмент также определяет пространственные выбросы. Для этого инструмент вычисляет локальное значение I Морана, z-оценку, псевдо-значение p и код, представляющий тип кластера для каждой статистически значимой функции.Z-значения и псевдо-p-значения представляют статистическую значимость вычисленных значений индекса. Просмотр дополнительных математических расчетов для местной статистики Морана I. Положительное значение для I указывает, что объект имеет соседние объекты с одинаковыми высокими или низкими значениями атрибутов; эта функция является частью кластера. Отрицательное значение для I указывает, что у объекта есть соседние объекты с разными значениями; эта особенность — выброс.В любом случае p-значение для признака должно быть достаточно маленьким, чтобы кластер или выброс считались статистически значимыми. Дополнительные сведения об определении статистической значимости см. В разделе Что такое z-оценка? Что такое p-значение? Обратите внимание, что местный индекс I Морана (I) является относительной мерой и может быть интерпретирован только в контексте его вычисленного z-значения или p-значения. Z-оценки и p-значения, указанные в выходном классе пространственных объектов, не корректируются для множественного тестирования или пространственной зависимости. Поле типа кластера / выброса (COType) различает статистически значимый кластер высоких значений (HH), кластер низких значений (LL), выброс, в котором высокое значение окружено преимущественно низкими значениями (HL), и выброс в котором низкое значение окружено преимущественно высокими значениями (LH). Статистическая значимость установлена на уровне достоверности 95 процентов. Если коррекция FDR не применяется, объекты с p-значениями меньше 0,05 считаются статистически значимыми.Коррекция FDR снижает этот порог p-значения с 0,05 до значения, которое лучше отражает 95-процентный уровень достоверности при многократном тестировании. Этот инструмент создает новый выходной класс пространственных объектов со следующими атрибутами для каждого объекта входного класса пространственных объектов: локальный индекс Морана I, z-оценка, p-значение и COType. При запуске этого инструмента выходной класс пространственных объектов автоматически добавляется в оглавление (TOC), а рендеринг по умолчанию применяется к полю COType.Применяемая визуализация определяется файлом слоя в Перестановки используются для определения вероятности нахождения фактического пространственного распределения значений, которые вы анализируете, путем сравнения ваших значений с набором случайно сгенерированных значений. Даже при полной пространственной случайности (CSR) некоторая степень кластеризации всегда будет наблюдаться просто из-за случайности.Перестановки будут генерировать множество случайных наборов данных и сравнивать эти значения с локальным индексом Морана I ваших исходных данных. Для этого каждая перестановка случайным образом переупорядочивает значения окрестностей вокруг каждого объекта и вычисляет значение Local Moran I для этих случайных данных. Глядя на распределение локальных значений I Морана, сгенерированных из перестановок, вы можете увидеть диапазон значений I локального Морана, которые могут быть разумно вызваны случайностью. Если в ваших данных есть статистически значимый пространственный образец, вы ожидаете, что значения I локального Морана, сгенерированные из перестановок, будут отображать меньшую кластеризацию, чем значение I локального Морана из ваших исходных данных.Затем вычисляется псевдо-p-значение, определяя долю локальных значений I Морана, сгенерированных из перестановок, которые отображают большую кластеризацию, чем ваши исходные данные. Если эта пропорция (псевдо-p-значение) мала (менее 0,05), вы можете сделать вывод, что ваши данные действительно отображают статистически значимую кластеризацию. Выбор количества перестановок — это баланс между точностью и увеличенным временем обработки. Увеличение количества перестановок увеличивает точность за счет увеличения диапазона возможных значений псевдо-p.Например, для 99 перестановок точность значения псевдо-p составляет 0,01, а для 999 перестановок точность составляет 0,001. Эти значения вычисляются путем деления единицы на количество перестановок плюс один: 1 / (1 + 99) и 1 / (1 + 999). При первом исследовании проблемы можно использовать меньшее количество перестановок, но лучше всего увеличить количество перестановок до максимально возможного числа для окончательных результатов. Инструмент кластерного анализа и анализа выбросов (Anselin Local Moran’s I) определяет концентрации высоких значений, концентрации низких значений и пространственные выбросы. Он может помочь вам ответить на такие вопросы, как эти: Приложения можно найти во многих областях, включая экономику, управление ресурсами, биогеографию, политическую географию и демографию. Анселин, Люк. «Местные индикаторы пространственной ассоциации — LISA», Географический анализ 27 (2): 93–115, 1995. Митчелл, Энди. Руководство ESRI по ГИС-анализу, Том 2. ESRI Press, 2005. Отзыв по этой теме? Независимо от того, являетесь ли вы аналитиком, черным поясом мастера бережливого производства и шести сигм, исследователем, студентом-статистиком или всеми вышеперечисленными, в какой-то момент вам понадобится работать с данными, которые включают даты, время или и то, и другое.Вы знаете, где находятся все ваши инструменты даты / времени? Давайте быстро познакомимся с программным обеспечением Minitab Statistical Software и посмотрим, что оно предлагает для данных даты и времени: Minitab позволяет легко управлять данными даты и времени и манипулировать ими без необходимости набирать ни одной строчки кода. Изменение формата даты или времени, извлечение элементов из столбца даты / времени или вычисление прошедшего времени — это легкое дело в статистическом программном обеспечении Minitab. Для иллюстрации я возьму случайную выборку из 100 звонков в колл-центр в период с 7 по 18 сентября.Данные упорядочены в таблице от самых старых до самых новых. Во-первых, нам может потребоваться вычислить прошедшее время, чтобы определить, сколько времени потребовалось оператору, чтобы ответить на каждый звонок, и продолжительность каждого звонка. Меню калькулятора Minitab предлагает мощный и полный каталог функций для управления числовыми, текстовыми данными или данными даты / времени. Истекшее время Функция идеально подходит для сценария нашего call-центра. Используйте Calc> Калькулятор или щелкните правой кнопкой мыши на листе и выберите Формулы> Назначить формулу столбцу : Установка флажка Назначить формулой или доступ к калькулятору путем щелчка правой кнопкой мыши на рабочем листе гарантирует, что вычисленное прошедшее время будет обновляться автоматически по мере добавления в рабочий лист большего количества вызовов. Истекшее время отлично подходит для визуализации изменения времени вызова на графике временного ряда. Затем функцию чистки Minitab можно использовать для определения продолжительного времени разговора и сосредоточения усилий по улучшению. Профессиональный совет : Чтобы уменьшить беспорядок на графике, я щелкнул правой кнопкой мыши по столбцу Получен вызов и выбрал столбец формата , чтобы удалить чч: мм: сс (время, указанное в таблице). Не волнуйтесь, это не удаляет раза, а только скрывает их на экране рабочего листа, чтобы они лучше смотрелись на графике.Время по-прежнему хранится в рабочем листе и может быть повторно отображено в окне «Формат столбца». Чтобы полностью удалить связанные времена, чтобы столбец даты можно было использовать в качестве группирующей переменной, используйте вместо этого Data> Date / Time> Round Date / Time . Чтобы визуализировать распределение общей продолжительности вызова в минутах, мы сначала используем Data> Date / Time> Extract to Numeric , чтобы извлечь только минуты из прошедшего времени. Логика транскрипции в случае частичной кооперативности между TF
Как работает анализ кластеров и выбросов (Anselin Local Moran’s I) — ArcGIS Pro
Расчеты
Интерпретация
Выходные данные
Перестановки
Рекомендации по передовой практике
Возможные приложения
Дополнительные ресурсы
Советы и приемы для данных даты / времени
Добавить комментарий